Сварка корня шва с обратным валиком | MastakSvarka
Материал создан при поддержке Ligans.
Особенностью моей работы является оценка начинающих сварщиков, выступающих на конкурсе по сварке. Мы часто с коллегами обсуждаем работы конкурсантов и спорим о том, что важнее в качестве сварного шва выполненного студентом. Первое, на что обращают внимание члены жюри, это внешний вид сварного шва. Я же считаю, что самое главное в сварном шве, это наличие качественного обратного валика. В горизонтальном положении начинающему сварщику не просто выполнить его, необходимо соблюдать три самых важных условия:
- ответственная подготовка деталей под сварку;
- правильно выбранный сварочный ток;
- качественные электроды
Очень важно правильно собрать детали под сварку соблюдая все допустимые размеры: зазор (2.5 мм), притупление кромок (1.5 — 2 мм).
Размеры зазора между деталями и притупление кромокВыбор сварочного тока произвожу на отдельной тренировочной детали, повторяющей форму сварного соединения ответственной детали.
Подбор сварочного токаСварку корня шва можно производить электродами диаметром 2.5 или 3 мм. Для сварки электродами Ligans SG 52TP диаметром 3 мм, я остановился на отметке сварочного тока 75 А.
Движение электродом выполняю «вперед- назад» слева на право, углом электрода назад. При движении вперед стараемся продавливать электрод в глубину зазора для того, чтобы образовывался качественный обратный валик.
При смене электрода необходимо зачистить место остановки ( замок) с помощью «Болгарки» , иначе в этом месте может образоваться непровар сварного шва.
Зачищенный замокЕсли правильно соблюдать эти три важных условия, то результат в виде сварного шва с обратным валиком приблизит вас к победе.
Обратная сторона сварного шваИ как всегда Вы можете посмотреть подробный ролик сварки горизонтального многопроходного сварного шва:
Методы сварки
Сварка нержавеющей стали
Сварка всех изделий, изготовленных из нержавеющей стали на заводе НПО ПМП Вентиляция, осуществляется аргонно-дуговым методом, с защитой «корня шва» инертным газом. Суть метода показана ниже.
Суть метода показана ниже.
На фото выше, представлена лицевая сторона сварного шва. Сваривали два листа нержавеющей стали толщиной 1,5 мм, сварка производилась аргонно-дуговым методом. Температура в зоне сварки доходит до 2000 ºС, что приводит к быстрому окислению расплавленного и разогретого металла при его взаимодействии с кислородом воздуха. Для предотвращения данного процесса в зону сварки подают инертный газ, в данном случае аргон. Аргон вытесняет воздух, и окисление не происходит. На фото хорошо видно, что в защитной среде аргона сталь практически не окисляется, шов выглядит ровным.
На фото выше, представлен тот же самый сварной шов, но после химической очистки (химического травления) в среде раствора кислот. В процессе химической очистки удаляются оксиды железа, образующиеся в процессе сварки в зоне шва, поверхность изделия очищается от попавших на нее, в процессе производства, соединений железа и органики.
На фото выше, мы видим обратную сторону сварного шва – его корень. Если лицевая сторона шва, там, где происходит сварка, обдувается инертным газом, то на другой стороне шва обдув отсутствует. Кислород воздуха легко вступает в химическую реакцию с сильно разогретым металлом, образуя неровный пористый шов с большим содержанием окислов.
На фото выше, это тот же корень шва, но после химической очистки. Очень хорошо видно, что даже длительное химическое травление не способно убрать все окислы из пористой структуры и очистить шов. В результате, это место имеет пониженную прочность и будет подвергаться коррозии. Кроме этого, в некоторых технологических процессах, с использованием труб и деталей из нержавеющей стали, наличие такого дефекта недопустимо.
Надо отметить, что некоторые производители для исключения такого дефекта, проваривают пластины с одной стороны не на полную глубину, сваривается только верхний слой пластин глубиной 0,5 – 1,0 мм. Результат очевиден – пониженная стойкость шва к механическим нагрузкам.
На фото выше, тоже представлена обратная сторона сварного шва (корень). Метод сварки тот же самый, толщина свариваемых листов нержавеющей стали 1,5 мм, но в процессе сварки, производился обдув обратной стороны шва инертным газом. В результате, мы имеем листы, сваренные по всей толщине, и отсутствие повреждений шва кислородом воздуха на корне.
На фото выше, тот же самый шов (корень), но после химической очистки. Корень шва чистый, не имеет выступов, края пластин сварены по всей толщине. Данный шов будет прочным и коррозионно стойким. На такую поверхность отлично ложатся полимерные покрытия, например тефлон и его производные.
Сварка углеродистой (черной) стали
Чаще всего углеродистую сталь сваривают полуавтоматическим способом с присадкой из нержавеющей стали и обдувом смесью углекислого газа и аргона. Этот метод хорош для крупных изделий с толщиной стенок от 3,0 мм и более. Корень шва не обдувается инертным газом, так как провар металла происходит не на всю толщину. Кроме этого, брызги расплавленного металла сильно портят внешний вид изделий. Для предотвращения данного эффекта, поверхность изделий, в месте сварки, предварительно покрывают специальной жидкостью.
На двух фотографиях выше, показаны сварные швы, выполненные полуавтоматическим методом. После механической очистки, поверхность окрашена. Швы сильно выделяются на поверхности изделия даже после механической обработки и покраски.
На фото выше, показан сварной внутренний угловой шов, выполненный аргонно-дуговым методом. Поверхность окрашена. Шов практически не заметен.
|

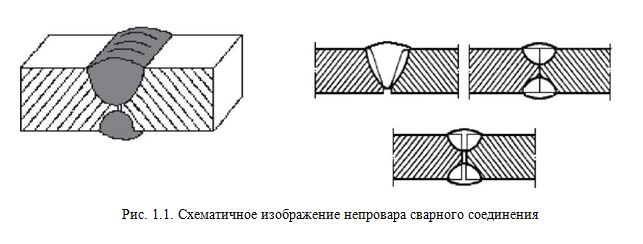
ПРИЕМЫ УДЕРЖАНИЯ МЕТАЛЛА. ПРИ НАЛОЖЕНИИ КОРНЕВОГО СЛОЯ ШВА
При выполнении корневого слоя шва основной трудностью является удержание расплавленного металла в сварочной ванне. Для предупреждения вытекания расплавленного металла из сварочной ванны применяют следующее:
стальные, медные, керамические или асбестовые подкладки, устанавливаемые под корень шва;
различного рода вставки между стыкуемыми кромками;
обходятся без сквозного проплавления с последующей подрубкой и подваркой корня шва с противоположной стороны;
флюсовые, флюсо-медные и газовые подушки, устанавливаемые под корень шва;
при стыковой сварке — соединения в замок;
специальные электроды, сварка которыми производится сверху вниз;
импульсную дугу.
Подкладки позволяют увеличивать величину сварочного тока, способствуя сквозному проплавлению свариваемых кромок шва и обеспечивая при этом 100% провар свариваемых деталей, а также задерживают и не позволяют расплавленному металлу вытекать из сварочной ванны и предупреждают образование прожогов. Для предупреждения вытекания расплавленного металла из сварочной ванны подкладки должны плотно прилегать к свариваемым кромкам.
Для предупреждения вытекания расплавленного металла из сварочной ванны подкладки должны плотно прилегать к свариваемым кромкам.
Установление различного рода вставок между стыкуемыми кромками способствует удержанию металла в сварочной ванне и 100% проплавлению свариваемых кромок в корне шва.
Иногда по заранее продуманной технологии не делают сквозного проплавления свариваемых кромок, а затем с обратной стороны делают канавку пневмозубилом или шлифмашинкой и накладывают подварочный шов, обеспечивая при этом 100%-ный провар свариваемых кромок в корне шва.
Расплавленный металл сварочной ванны в корне шва удерживают применением различных подушек. Флюсовые и флюсо-медные подушки применяются при сварке под флюсом. Газовая подушка используется при ручной дуговой сварке, ручной и автоматической аргонодуговой сварке с использованием в качестве поддувочного газа аргона, азота или углекислого газа. Сущность газовой подушки состоит в том, чго под расплавленный металл сварочной ванны подается один из выше упомянутых газов с избыточным давлением 50—200 мм вод. ст.
Соединение в замок за счет металла одной из стыкуемых кромок позволяет избежать прожогов в корне шва.
При сварке сверху вниз специальными электродами дюкрытие электрода приводит к увеличению силы поверхностного натяжения металла, в результате чего Исключаются прожоги в корне шва.
Импульсная дуга приводит к кратковременному расплавлению металла, в результате чего металл корня сварного шва охлаждается и кристаллизуется. На остывающую порцию расплавленного металла накладывается новая, а так как дуга на некоторый промежуток времени прерывается и новый металл в это время не добавляется, то и не будет прожогов.
Системы водоснабжения (стр. 5 из 7)
Движения «полумесяцем назад» используют для сварки в нижнем положении, а также для вертикальных и потолочных швов с выпуклой наружной поверхностью.
Движения «треугольником» применяют для угловых швов с катетом более 6мм и стыковых швов со скосом кромок в любом пространственном положении. Даёт хороший провар корня шва.
Даёт хороший провар корня шва.
Движения «треугольником» с задержкой электрода в корне шва эффективны для сварки толстостенных конструкций с гарантированным проплавлением корневого участка шва.
Петлеобразные движения используют для усиленного прогревания кромок шва, особенно при сварке высоколегированных сталей. Электрод задерживают на краях, чтобы не было прожога в центре шва или вытекания металла при сварке вертикальных швов.
Выполнение швов в различных положениях.
Выполнение стыковых соединений в нижнем положении. Наиболее удобно выполнять сварку в нижнем положении, швы получаются высокого качества, так как в этом случае легко выделяются неметаллические включения и газы из расплавленного металла сварочной ванны. При этом также имеются лучшие условия для формирования металла шва, поскольку расплавленный металл сварочной ванны удерживается от вытекания нерасплавившимися кромками.
Наложение валиков рекомендуется производить слева направо или к себе. В этих случаях сварщик чётко видит место соединения, длину дуги, перенос капель электродного металла и формирование валика. Нормальной считается ширина валика, равная 3–4 диаметрам электрода.
Односторонние швы без скоса кромок выполняют электродами диаметром, равным толщине металла S, если она превышает 4мм.
Листы без скоса кромок толщиной 2–8мм сваривают двусторонним швом, а до 6мм – односторонним.
Металл толщиной более 8мм сваривают с разделкой кромок. Во избежание прожогов сварку ведут на съемных медных или стальных подкладках.
Однопроходную сварку с V-образной разделкой кромок обычно выполняют поперечными колебаниями электрода на всю ширину, чтобы дуга перемещалась со скоса кромок на необработанную поверхность металла. Однако в этом случае очень трудно обеспечить равномерный провар шва по всей его длине, особенно при изменении величины притупления кромок и зазора между ними.
При сварке шва с V-образной разделкой за несколько проходов обеспечить хороший провар первого слоя в корне разделки гораздо легче. Для этого обычно применяют электроды диаметром 3-4мм и сварку ведут без поперечных колебаний. Последующие слои выполняют электродом большего диаметра (в соответствии с толщиной металла) с поперечными колебаниями. Для обеспечения хорошего провара между слоями предыдущие швы, а также кромки следует тщательно очищать от шлака и брызг металла.
Для этого обычно применяют электроды диаметром 3-4мм и сварку ведут без поперечных колебаний. Последующие слои выполняют электродом большего диаметра (в соответствии с толщиной металла) с поперечными колебаниями. Для обеспечения хорошего провара между слоями предыдущие швы, а также кромки следует тщательно очищать от шлака и брызг металла.
Сварку швов с X- или U-образной разделкой кромок выполняют так же, как и с V-образной. Однако для уменьшения остаточных деформаций и напряжений сварку ведут, накладывая каждый последующий шов навстречу предыдущему. Швы с Х-образным скосом кромок более предпочтительны, чем с V-образным, так как в 1,6-1,7 раза уменьшается объем наплавленного металла, повышается производительность сварки и, кроме того, снижается величина угловых деформаций.
Сварку стыковых швов можно выполнять различными способами. При сварке на весу трудно обеспечить провар корня шва и формирование хорошего обратного валика по всей длине стыка. Поэтому используют сварку на съемной медной или остающейся стальной подкладке. В медной подкладке для формирования обратного валика делают формирующую канавку. Для того чтобы предотвратить вытекание расплавленного метала из сварочной ванны, необходимо плотно поджимать подкладки к свариваемым кромкам.
Если с обратной стороны соединения возможен подход к корню шва и допустима выпуклость обратной стороны шва, целесообразно выполнить подварку корня швом небольшого сечения с последующей укладкой основного шва с лицевой стороны соединения.
В местах поворота сварной шов следует заваривать без отрыва дуги. Не допускается гашение и зажигание дуги на поворотах сварного шва.
Выполнение угловых соединений в нижнем положении.
Выполняют угловые соединения «в симметричную лодочку» (рис.20,а) и «в несимметричную лодочку» (рис.20,б)
Во избежание непровара и подрезов кромок сварку «в лодочку» лучше вести электродом, допускающим опирание покрытия (козырька) на кромки.
При наложении угловых швов наклонным электродом (в том числе «в лодочку») сварку лучше вести «углом назад».
При выполнении тавровых соединений дугу возбуждают на горизонтальной полке, а не на вертикальной, чтобы избежать натекания металла.
Угловые швы без скоса кромок с катетами более 10мм выполняют в один слой поперечными движениями электрода «треугольником» с задержкой в корне шва.
Сварка угловых швов нахлесточных соединений в нижнем положении с катетом до 10мм производится в один слой электродами диаметром до 5мм без поперечных колебаний.
Выполнение вертикальных швов.
Сварка швов в положениях, отличающихся от нижнего, требует высокой квалификации сварщика. При ее выполнении возможны вытекание расплавленного металла из сварочной ванны или падение капель электродного металла мимо нее. Для предотвращения этого сварку следует вести по возможности наиболее короткой дугой, в большинстве случаев с поперечными колебаниями.
Расплавленный металл в сварочной ванне удерживается от вытекания силой поверхностного натяжения, поэтому необходимо уменьшить ее объем. Для этого конец электрода периодически отводят в сторону от ванны, давая возможность ей частично закристаллизоваться. Ширину валиков также уменьшают до двух-трех диаметров электрода. Применяют электроды меньших диаметров (для вертикальных и горизонтальных швов – не более 5мм, для потолочных – не более 4мм).
Выполняя вертикальные швы, силу сварочного тока уменьшают на 10% по сравнению со сваркой в нижнем положении. Для того чтобы металл не вытекал из ванны, нужно поддерживать короткую дугу. Используются электроды, дающие быстротвердеющий тонкий слой шлака («короткие» шлаки).
При способе «снизу вверх» («на подъем») дугу возбуждают в нижней точке шва. Сваркой подготавливают горизонтальную площадку сечением, равным сечению шва. При этом электрод совершает поперечные колебания. Сварка этим способом обеспечивает возможность провара корня шва и кромок, так как расплавленный металл стекает с них в сварочную ванну, улучшая условия теплопередачи от дуги к основному металлу. Однако поверхность шва получается грубочешуйчатая.
Наибольший провар достигается при положении электрода, перпендикулярном вертикальной оси. Стекание расплавленного металла предотвращают наклоном электрода вниз.
Сварка «на подъем»- наиболее удобный, распространенный способ. Используются электроды диаметром до 4мм. Поперечные колебания электрода: «углом», «полумесяцем», «елочкой».
При способе «сверху вниз» («на спуск») дугу возбуждают в верхней точке шва. После образования капли жидкого металла электрод наклоняют так, чтобы дуга была направлена на жидкий металл.
Сварка «на спуск» затрудняет получение качественного провара: шлак и расплавленный металл подтекают под дугу и от дальнейшего стекания удерживаются только силами давления дуги и поверхностного натяжения. Иногда их оказывается недостаточно, и расплавленный металл вытекает из сварочной ванны.
Сварка «сверху вниз» позволяет избежать прожогов при соединении тонкого металла. Рекомендуется в основном для сварки тонких (до 5мм) листов с разделкой кромок. Используются электроды с целлюлозным покрытием (марок ОЗС-9, АНО-9, ВСЦ-2, ВСЦ-3).
При сварке «по спирали» или «полумесяцем» вначале направляют полочку на свариваемые кромки, а затем небольшими порциями направляют металл, постепенно перемещая электрод выше, оставляя внизу готовый сварной шов.
При сварке «углом» электрод попеременно перемещают вверх-вниз, беспрерывно наплавляя металл на кромки и равномерно перенося его вверх электродом.
При сварке «елочкой» вначале электрод поднимают вверх вправо, а затем опускают вниз. Капля жидкого металла застывает между кромками. Затем электрод поднимают, вверх вправо и снова опускают вниз, оставляя новую порцию металла.
Выполнение горизонтальных швов. Сварка горизонтальных стыковых швов более затруднительна, чем вертикальных, из-за стекания расплавленного металла из сварочной ванны на нижнюю кромку. В результате возможно образование подреза по верхней кромке. При сварке горизонтальных стыковых швов необходим скос только верхней кромки. Дугу возбуждают на нижней (рис.24) горизонтальной кромке, а затем электрод переводят на верхнюю.
Дугу возбуждают на нижней (рис.24) горизонтальной кромке, а затем электрод переводят на верхнюю.
Сварку горизонтальных стыковых швов можно вести вертикально расположенным электродом, а также «углом вперед» и «углом назад» (рис.25).
Очередность выполнения проходов при сварке горизонтальных стыковых швов показана на рисунке 26.
Выполнение потолочных швов. Сварка таких швов наиболее сложна. Газы, выделяемые покрытием электрода, поднимаются вверх и могут остаться в шве, поэтому используют хорошо просушенные электроды. Удлинение дуги нередко вызывает образование подрезов. Узкие валики накладывают в разделку тремя способами.
При сварке «лесенкой» электрод располагают под углом к плоскости 90-130о, подводят к изделию и зажигают дугу. После образования маленькой порции расплавленного металла электрод отводят на 5-10мм от потолочной плоскости и возвращают, перекрывая закристаллизовавшуюся порцию металла расплавленным примерно на 1/2 – 1/3 ее длины (рис.27,а).
При сварке «полумесяцем» электрод располагают под углом 90-1300 к потолочной плоскости и, манипулируя по схеме полумесяца, беспрерывно заходят электродом на закристаллизовавшуюся часть металла (рис.27,б).
Автоматическая сварка под флюсом
Последние годы на заводах находит применение автоматическая сварка под флюсом с применением порошкообразного присадочного металла (ППМ). Этот вид сварки по сравнению с обычным способом автоматической сварки под флюсом имеет следующие преимущества:
скоpость сварки стыковых и угловых соединений увеличивается в 1,5—2 раза;
сварка стыковых соединений толщиной до 60 мм выполняется по одному проходу с каждой стороны без разделки кромок;
механические свойства сварного шва улучшаются за счет уменьшения перегрева металла сварочной ванны и кромок изделия;
уменьшается расход флюса в 1,5 раза;
снижается себестоимость в 2 раза.
В качестве ППМ применяют железный порошок ПЖ-2К по ГОСТ 9849—74* и крупку из электродной проволоки диаметром 0,8 1,6 мм, нарубленную длиной не более диаметра проволоки. При диаметре проволоки 2 мм длина крупки должна быть не более 1 мм.
При диаметре проволоки 2 мм длина крупки должна быть не более 1 мм.
Электрошлаковая сварка позволяет полностью отказаться от пакетов, заменив их толстыми листами в сплошностенчатых конструкциях. Основным преимуществом электрошлаковой сварки является возможность сварки деталей практически неограниченной толщины в один слой без обработки кромок. Вследствие симметричного расположения свариваемых кромок отсутствуют угловые деформации от сварки. Электрошлаковую сварку обычно применяют для сварки деталей толщиной свыше 45 мм.
Порядок наложения швов зависит от вида конструкций и способов сварки. Порядок наложения швов должен обеспечивать высокое качество сварных соединений, максимальную производительность, минимальные сварочные деформации, удобство и безопасность работ. Сварку конструкций необходимо начинать со стыков, которые следует сваривать предварительно в нижнем положении, преимущественно механизированными способами. При сварке стыковых швов должны быть обеспечены полный провар, отсутствие внутренних дефектов и хороший внешний вид. Перед наложением второго шва необходимо из зазора между свариваемыми деталями удалить шлак и грат с внутренней части первого (подварочного) шва путем вырубки корня шва. Это обеспечивает полный провар, отсутствие внутренних дефектов и проектную прочность шва.
Удаление корня шва производится абразивными армированными кругами толщиной 3—6 мм, пневматическим зубилом или воздушно-дуговым способом с последующей зачисткой стальными щетками. При воздушно-дуговом способе между корнем шва и угольным электродом возбуждается электрическая дуга, расплавляется часть основного и наплавленного металла, который вместе со шлаком выдувается сжатым воздухом.
Полный провар стыковых швов, выполняемых автоматами, обеспечивается односторонней сваркой на флюсовой подушке или стальной подкладке, а также двухсторонней сваркой. Флюсовая подушка обеспечивает полный провар при односторонней сварке автоматами, металла толщиной до 10 мм без разделки кромок и до 24 мм — с V-образной разделкой.
Термины, описывающие процессы сплавления и проплавления. |
Понятие «сплавление» относится, как правило, к фактическому совместному плавлению присадочного и основного металлов или к плавлению только основного металла. Понятие «проплавление» относится к расстоянию, на которое наплавленный металл проникает в стык. Степень достигнутого проникновения непосредственно влияет на прочность соединения и поэтому принадлежит к размерным параметрам шва.
Для описания степени и места сплавления или проплавления имеются многочисленные понятия. Поскольку в процессе сварки происходит плавление первоначальной поверхности разделки, конечные границы металла сварного шва находятся глубже, чем первоначальные поверхности. Поскольку поверхность разделки плавится в процессе сварки, в дальнейшем она называется свариваемой кромкой. Граница перехода между наплавленным металлом и основным металлом называется границей раздела сварного шва. Глубина проплавления – это «расстояние от свариваемой кромки до границы раздела сварного шва». Глубина проплавления всегда измеряется в направлении нормали к поверхности контакта между свариваемыми элементами. Зона проплавления – это «область расплавленного основного металла, определенная на поперечном сечении сварного шва». Эти же термины применяются подобным образом и другим видам швов, например, угловым швам и наплавке поверхности. Иллюстрацией к понятиям, связанным с плавлением, может служить рис.1.
Как показано на рис.2, несколько понятий используются также для параметров проплавления в сварном шве. Проплавление (провар) корня шва – это «расстояние, на которое расплавленный металл проникает за пределы зазора между свариваемыми кромками». Глубина проплавления стыка – это «расстояние, на которое расплавленный металл проникает от наиболее удаленной точки лицевой стороны сварного шва в стык, за вычетом значения возможной выпуклости шва». У швов с разделкой кромок это же самое расстояние называется также размером сварного шва.
Еще одно смежное понятие – это зона термического влияния. Эта область, показанная на рис.3, определяется как «та часть основного металла, которая не расплавилась, но механические свойства или микроструктура которой изменились из-за нагрева во время сварки, пайки твердым припоем, пайки мягким припоем или резки».
Проникновение корней в глубокие слои почвы стимулирует минерализацию тысячелетнего органического углерода
Основные моменты
- •
Уязвимость глубокого углерода к минерализации была оценена в эксперименте по мезокосму.
- •
Глубоко укоренившаяся трава выращивалась на почвенных столбах с двойной маркировкой ( 13 C и 14 C-CO 2 ).
- •
Глубокие корни стимулировали микробную минерализацию возрастом ~ 15 000 лет C.
- •
Deep C так же уязвим для минерализации, вызванной ризосферой, как и поверхность C.
Abstract
Изменения климата и землепользования изменяют глубину укоренения растений, что означает, что органическое вещество с длительным временем пребывания в глубоких слоях слои почвы могут подвергаться воздействию ризосфер и связанной с ними микробной активности. Присутствие корней в почвах стимулирует минерализацию естественного почвенного углерода за счет процесса, называемого эффектом ризосферы ризосферы (RPE), который, как следствие, может привести к потере почвенного C.Выращивая траву с глубокими корнями, Festuca arundinacea , на столбиках почвы и при непрерывной двойной маркировке ( 13 C- и 14 C-CO 2 ), мы показали, что корни проникают в почву на глубину до 80 см. профиль стимулировал минерализацию почвы C. возрастом около 15000 лет. RPE после нормализации с корневой биомассой был аналогичным по профилю почвы, что указывает на то, что глубокий C так же уязвим для грунтования, как и поверхностный C.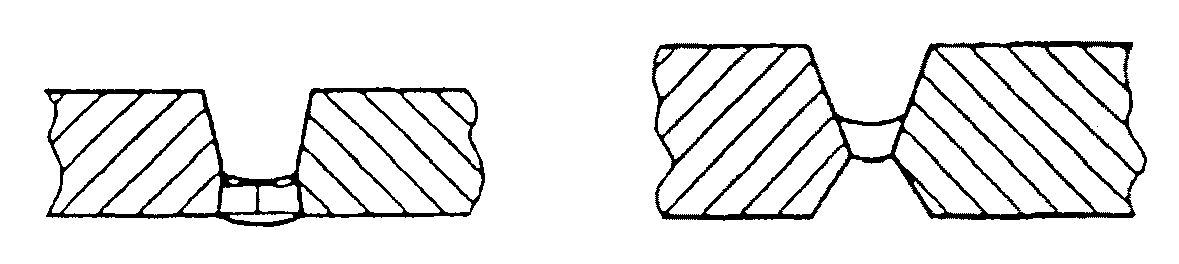 RPE сильно коррелировал с дыханием растений. производное C и маркер PLFA, представляющий сапрофитные грибы (18: 2-6c) во всех слоях почвы.Более того, экспериментальное нарушение структуры почвы еще больше стимулировало минерализацию почвы углеродом. Эти данные свидетельствуют о том, что медленная минерализация почвенного углерода в глубоких слоях является результатом обеднения богатого энергией растения С для микроорганизмов (особенно для сапрофитных грибов) в сочетании с физическим разрывом между почвенным углеродом и микроорганизмами. Основываясь на наших результатах, мы ожидаем более высоких темпов минерализации глубокого тысячелетнего ПОВ в ответ на более глубокое проникновение корней, которое может быть вызвано изменениями в методах ведения сельского хозяйства и климате.
RPE сильно коррелировал с дыханием растений. производное C и маркер PLFA, представляющий сапрофитные грибы (18: 2-6c) во всех слоях почвы.Более того, экспериментальное нарушение структуры почвы еще больше стимулировало минерализацию почвы углеродом. Эти данные свидетельствуют о том, что медленная минерализация почвенного углерода в глубоких слоях является результатом обеднения богатого энергией растения С для микроорганизмов (особенно для сапрофитных грибов) в сочетании с физическим разрывом между почвенным углеродом и микроорганизмами. Основываясь на наших результатах, мы ожидаем более высоких темпов минерализации глубокого тысячелетнего ПОВ в ответ на более глубокое проникновение корней, которое может быть вызвано изменениями в методах ведения сельского хозяйства и климате.
Ключевые слова
Глубинный углерод почвы
Эффект прайминга ризосферы
14 C-датирование
13 C-маркировка
Структура микробного сообщества
Ограничение энергии микробов
Рекомендуемая стойкость почвы
C 9000 статей Просмотреть полный текст© 2018 Elsevier Ltd. Все права защищены.
Рекомендуемые статьи
Цитирующие статьи
Проникновение корней через слой почвы с высокой насыпной плотностью: дифференциальная реакция растений и сорняков
Bengough AG, Mullins CE (1990) Механическое сопротивление росту корней: обзор экспериментальных методов и реакции роста корней.J Soil Sci 41: 341–358
Google Scholar
Bennie ATP (1996) Рост и механическое сопротивление. В: Waisel Y, Eshel A, Kafkafi U (ред.) Корни растений: скрытая половина. Dekker Publishers, Нью-Йорк, стр. 453–470
Google Scholar
Бенш К.Н., Хорак М.Дж., Петерсон Д. (2003) Взаимодействие краснокорневых свиных водорослей, амаранта Палмера и водяной конопли в сое. Weed Sci 51: 37–43
Статья CAS Google Scholar
Buol SW, Southard RJ, Graham RC, McDaniel PA (2003) Генезис и классификация почв, 5-е изд. Iowa State University Press, Ames, pp. 1–494
Iowa State University Press, Ames, pp. 1–494
Бушамука В.Н., Зобель Р.В. (1998) Дифференциальное генотипическое и корневое проникновение в уплотненные слои почвы. Crop Sci 38: 776–781
Google Scholar
Busscher WJ, Lipiec J, Bauer PJ, Carter TE Jr (2000) Улучшенное проникновение корней в твердые слои почвы благодаря выбранному генотипу. Commun Soil Sci Plant Anal 31: 3089–3101
CAS Google Scholar
Campbell RB, Reicosky DC, Doty CW (1974) Физические свойства и обработка палеодультов на юго-восточных прибрежных равнинах.J Soil Water Conserv 29: 220–224
Google Scholar
Campbell BD, Grime JP, Mackey JML (1991) Компромисс между масштабом и точностью при поиске ресурсов. Oecologia 87: 532–538
Статья Google Scholar
Carmi A, Heuer B (1981) Роль корней в контроле роста побегов фасоли. Энн Бот 48: 519–527
CAS Google Scholar
Carter TE Jr, Rufty TW (1993) Интродукция растений сои, демонстрирующих засухоустойчивость и устойчивость к алюминию.В: Адаптация пищевых культур к температуре и водному стрессу: материалы международного симпозиума, Тайвань. Азиатский центр исследований и разработок овощей, Шанхуа, Тайвань, стр. 335–346
Champoux MC, Wang G, Sarkarung S, Mackill DJ, O’Toole JC, Huang N, McCouch SR (1995) Поиск генов, связанных с морфологией корня и предотвращение засухи риса за счет связи с молекулярными маркерами. Theor Appl Genet 90: 969–981
Статья CAS Google Scholar
Cook A, Marriott CA, Seel W, Mullins CE (1996) Влияние механического сопротивления почвы на рост корней и побегов Lolium perenne L., Капилляры Agrostis и Trifolium repens L.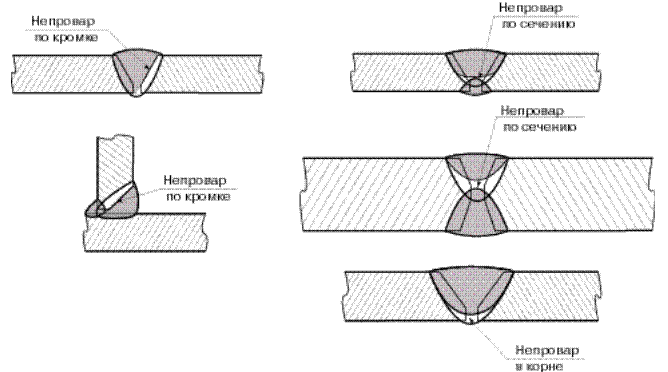 J Exp Bot 47: 1075–1084
J Exp Bot 47: 1075–1084
Артикул CAS Google Scholar
Crick JC, Grime JP (1987) Морфологическая пластичность и захват минеральных питательных веществ у двух травянистых видов с контрастирующей экологией. New Phytol 107: 403–414
Артикул Google Scholar
Culpepper AS, Gray TL, Vencill WK, Kichler JM, Webster TM, Brown SM, York AC, Davis JW, Hanna WW (2006) Устойчивый к глифосату амарант Палмера ( Amaranthus palmeri ) подтвержден в Джорджии.Weed Sci 54: 620–626
Статья CAS Google Scholar
Дрю М.К., балобан Л.Р. (1975) Обеспечение питательными веществами и рост семенной корневой системы ячменя. II. Локальное компенсаторное увеличение бокового роста корней и скорости поглощения нитратов, когда подача нитратов ограничена только частью корневой системы. J Exp Bot 26: 79–90
Статья CAS Google Scholar
Дунбабин В. (2007) Моделирование роли признаков укоренения в конкуренции посевов и сорняков.Полевые культуры Res 104: 44–51
Статья Google Scholar
Фельдман Л.Дж. (1984) Регулирование развития корневой системы. Annu Rev Plant Physiol 35: 223–242
PubMed Статья CAS Google Scholar
Монтажник А.Х. (1994) Архитектура и распределение биомассы как компоненты пластической реакции корневой системы на неоднородность почвы. В: Caldwell MM, Pearcy RW (eds) Использование растениями неоднородности окружающей среды.Academic, Сан-Диего, стр 305–323
Google Scholar
Foy CD (1984) Физиологические эффекты токсичности водорода, алюминия и марганца в кислой почве. В: Адамс Ф. (ред.) Кислотность почвы и известкование. Американское общество агрономии, Мэдисон, стр. 57–98
Американское общество агрономии, Мэдисон, стр. 57–98
Google Scholar
Фридман М., Хеника П.Р. (1991) Мутагенность токсичных семян сорняков в тесте Эймса: дурман ( Datura stramonium ), вельветлиф ( Abutilon theophrasti ), ипомея ( Ipomoea spp.) и серподы ( Cassia obtusifolia ). J Agric Food Chem 39: 494–501
Статья CAS Google Scholar
Гент Дж. А., Баллард Р., Хассан А. Е., Кассель Д. К. (1984) Влияние лесозаготовок и подготовки участка на физические свойства лесных почв Пьемонта. Soil Sci Soc Am J 48: 173–177
Google Scholar
Granato TC, Raper CD (1989) Разрастание корней кукурузы в ответ на локализованное поступление нитратов.J Exp Bot 40: 263–275
PubMed Статья CAS Google Scholar
Grime JP (1994) Роль пластичности в использовании неоднородности окружающей среды. В: Caldwell MM, Pearcy RW (eds) Использование растениями неоднородности окружающей среды. Academic, Сан-Диего, стр. 1–19
Google Scholar
Grime JP, Crick JC, Rincon JE (1986) Экологическое значение пластичности.В: Дженнингс Д.Х., Тревавас А.Дж. (ред.) Пластичность растений. Биологи Лимитед, Кембридж, стр. 4–29
Google Scholar
Harry-O’Kuru RE, Wu YV, Evangelista R, Vaughn SF, Rayford W., Wilson RF (2005) Обработка семян серподы ( Senna obtusifolia ) и потенциальное использование. J Agric Food Chem 53: 4784–4787
PubMed Статья CAS Google Scholar
Джексон Р.Б., Колдуэлл М.М. (1989) Сроки и степень разрастания корней на микросайтах плодородной почвы для трех многолетних растений холодных пустынь.Oecologia 81: 149–153
Google Scholar
Джексон Л.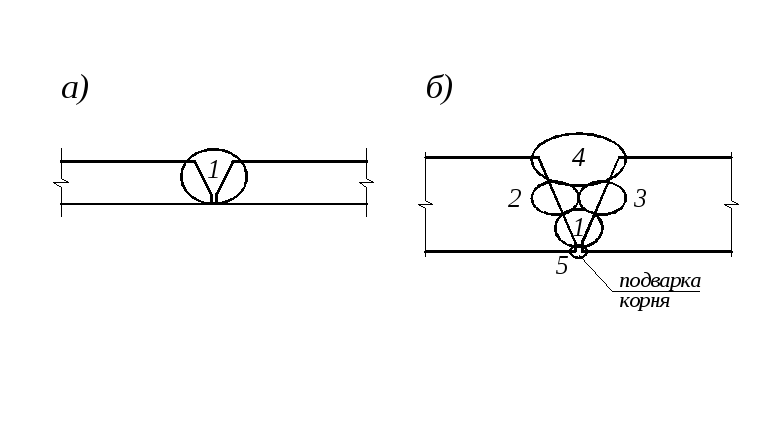 Е., Кох Г. В. (1997) Экофизиология сельскохозяйственных культур и их диких родственников. В: Джексон Л. Е. (ред) Экология в сельском хозяйстве. Academic, Сан-Диего, стр. 3–37
Е., Кох Г. В. (1997) Экофизиология сельскохозяйственных культур и их диких родственников. В: Джексон Л. Е. (ред) Экология в сельском хозяйстве. Academic, Сан-Диего, стр. 3–37
Google Scholar
Jackson RB, Manwaring JH, Caldwell MM (1990) Быстрая физиологическая адаптация корней к локальному обогащению почвы. Nature 344: 58–60
PubMed Статья CAS Google Scholar
Johnson WC, Jackson LE, Ochoa O, van Wijk R, Peleman J, St Clair DA, Michelmore RW (2000) Салат с мелкой корневой системой и Lactuca serriola , его дикий предок, различаются в QTL. определение корневой архитектуры и эксплуатации глубоких почвенных вод.Theor Appl Genet 101: 1066–1073
Статья CAS Google Scholar
Jones RE Jr, Walker RH, Wehtje G (1997) Соя ( Glycine max ), дурня обыкновенный ( Xanthium strumarium ) и серпод ( Senna obtusifolia ) в межвидовой конкуренции. Weed Sci 45: 409–413
CAS Google Scholar
Kasperbauer MJ, Busscher WJ (1991) Генотипические различия в проникновении корней хлопчатника в уплотненный слой почвы.Crop Sci 31: 1376–1378
Google Scholar
Keeley PE, Carter CH, Thullen RJ (1987) Влияние даты растения на рост амаранта Палмера. Weed Sci 35: 199–204
Google Scholar
Кембель С.В., Кэхилл Дж.Ф. (2005) Подземная фенотипическая пластичность растений: филогенетическая перспектива компромиссов при кормлении корнями. Am Nat 166: 216–230
PubMed Статья Google Scholar
Клингаман Т.Э., Оливер Л.Р. (1994) Палмер амарант ( Amaranthus palmeri ) вмешивается в соевые бобы ( Глицин макс ). Weed Sci 42: 523–527
Weed Sci 42: 523–527
CAS Google Scholar
Кубо К., Джитсуяма К., Ивама К., Хасегава Т., Ватанабе Н. (2004) Генотипические различия в способности проникать в корни твердой пшеницы ( Triticum turgidum L. var. durum ), оцениваемой в горшке с парафином. Вазелиновые диски. Растительная почва 262: 169–177
Артикул CAS Google Scholar
Lazof DB, Rufty TW, Redinbaugh MG (1992) Локализация абсорбции и транслокации нитратов в морфологических областях корня кукурузы.Plant Physiol 100: 1251–1258
PubMed CAS Статья Google Scholar
Масле Дж. (1992) Генетические вариации влияния импеданса корней на рост и скорость транспирации пшеницы и ячменя. Aust J Plant Physiol 19: 109–125
Google Scholar
Масле Дж., Пассиура Дж. Б. (1987) Влияние прочности почвы на рост молодых растений пшеницы. Aust J Plant Physiol 14: 643–656
Статья Google Scholar
Масле Дж., Фаркуар Г.Д., Гиффорд Р.М. (1990) Рост и углеродная экономия проростков пшеницы в зависимости от сопротивления почвы проникновению и парциального давления окружающей среды CO 2 .Aust J Plant Physiol 17: 465–487
CAS Google Scholar
Матеречера С.А., Декстер А.Р., Алстон А.М. (1991) Проникновение очень сильных почв корнями проростков различных видов растений. Растительная почва 135: 31–41
Статья Google Scholar
Матеречера С.А., Алстон А.М., Кирби Дж. М., Декстер А. Р. (1992) Влияние диаметра корня на проникновение семенных корней в уплотненный грунт.Растительная почва 144: 297–303
Артикул Google Scholar
Май О. Л., Каспербауэр М.Дж. (1999) Генотипические вариации проникновения корней в почвенный поддон. J Sustain Agric 13: 87–94
Л., Каспербауэр М.Дж. (1999) Генотипические вариации проникновения корней в почвенный поддон. J Sustain Agric 13: 87–94
Статья Google Scholar
Монкс Д.В., Оливер Л.Р. (1988) Взаимодействие между сортами сои ( Glycine max ) и отдельными сорняками. Weed Sci 36: 770–774
Google Scholar
Мойер-Генри К.А., Бертон Дж.М., Израиль Д.В., Руфти Т.В. (2006) Перенос азота между растениями: исследование естественного содержания 15 N среди сельскохозяйственных культур и сорняков.Растительная почва 282: 7–20
Артикул CAS Google Scholar
Naderman GC (1990) Уплотнение и углубление недр в Северной Каролине, обзор. Служба распространения сельскохозяйственных знаний Северной Каролины, публикация AG-353
Национальная служба сельскохозяйственной статистики [NASS] (2006) Выпущено в мае 2007 г. По состоянию на 2 марта 2008 г. (http://usda.mannlib.cornell.edu)
Nice GRW, Buehring NW, Shaw DR (2001) Sicklepod ( Senna obtusifolia ) реакция на затенение, междурядье сои ( Glycine max ) и численность в трех системах управления.Weed Technol 15: 155–162
Артикул Google Scholar
Норсуорси Дж.К. (2004) Влияние формирования растительного покрова сои на вылет без ямок ипомеи (Ipomoea lacunosa), дурничего обыкновенного (Xanthium strumarium) и серпопод (Senna obtusifolia). Weed Sci 52: 954–960
Статья CAS Google Scholar
Passioura JB (1991) Структура почвы и рост растений.Aust J Soil Res 29: 717–728
Статья Google Scholar
Pearson RW (1974) Значение укоренения для растениеводства и некоторые проблемы исследования корней. В: Carson EW (ed) Корень растения и его окружающая среда. Издательство Университета Вирджинии, Шарлоттсвилль, стр. 247–270
Издательство Университета Вирджинии, Шарлоттсвилль, стр. 247–270
Google Scholar
Price AH, Steele KA, Moore BJ, Barraclough PB, Clark LJ (2000) Комбинированная карта связей RFLP и AFLP риса с возвышенностей ( Oryza sativa L.), используемый для идентификации QTL для способности проникновения в корень. Theor Appl Genet 100: 49–56
Статья CAS Google Scholar
Радосевич С., Холт Дж, Герса С. (1997) Физиологические аспекты конкуренции (Глава 6). В кн .: Экология сорняков, 2-е изд. Wiley, New York, pp. 217–299
Ray JD, Yu L, McCouch SR, Champoux MG, Wang G, Nguyen HT (1996) Картирование локусов количественных признаков, связанных со способностью проникновения корней в рис.Theor Appl Genet 92: 627–636
Статья CAS Google Scholar
Робертс Дж. А., Хусейн А., Тейлор И. Б., Колин Р., Блэк CR (2002) Использование мутантов для изучения передачи сигналов на большие расстояния в ответ на уплотненную почву. J Exp Bot 53: 45–50
PubMed Статья CAS Google Scholar
Робинсон Д. (1994) Реакция растений на неравномерное поступление питательных веществ.New Phytol 127: 635–674
Артикул CAS Google Scholar
Рассел Р.С. (1977) Корневые системы растений: их функции и взаимодействие с почвой. Макгроу-Хилл, Англия, Великобритания
Google Scholar
Рассел Р.С., Госс М.Дж. (1974) Физические аспекты плодородия почвы — реакция корней на механическое сопротивление. Neth J Agric Sci 22: 305–318
Google Scholar
Schlichting CD (1986) Эволюция фенотипической пластичности у растений.Ann Rev Ecolog Syst 17: 667–693
Статья Google Scholar
Sellers BA, Smeda RJ, Johnson WG, Kendig JA, Ellersieck MR (2003) Сравнительный рост шести видов амаранта в Миссури. Weed Sci 51: 329–333
Weed Sci 51: 329–333
Статья CAS Google Scholar
Тейлор Х.М. (1974) Поведение корней в зависимости от структуры и прочности почвы. В: Carson EW (ed) Корень растения и его окружающая среда.Издательство Университета Вирджинии, Шарлоттсвилл, стр. 271–290
Google Scholar
Thomas JF, Downs RJ, Saravitz CH (2006) Руководство по процедурам фитотрона. Государственный университет Северной Каролины. Технический бюллетень 244
Унгер П.В., Каспар Т.К. (1994) Уплотнение почвы и рост корней: обзор. Agron J 86: 759–766
Google Scholar
Фон Икскалль Х., Мутерт Э. (1995) Глобальные масштабы, развитие и экономическое влияние кислых почв.Растительная почва 171: 1–15
Артикул Google Scholar
Webster TM (2005) Обследование сорняков — южные штаты: подраздел широколиственных культур. Proc South Weed Sci Soc 58: 291–304
Google Scholar
Webster TM, MacDonald GE (2001) Исследование сорняков на различных культурах в Джорджии. Weed Technol 15: 771–790
Статья Google Scholar
Wilcox JR (2004) Мировое распределение сои и торговля ею.В: Boerma HR, Specht JE (ред.) Соя: улучшение, производство и использование. Американское агрономическое общество, Мэдисон, Висконсин, стр. 1–14
Google Scholar
Wright SR, Jennette MW, Coble HD, Rufty TW (1999) Морфология корней молодых Glycine max , Senna obtusifolia и Amaranthus palmeri . Weed Sci 47: 706–711
CAS Google Scholar
Зимдал Р.Л. (1999) Основы науки о сорняках.Академик, Нью-Йорк
Google Scholar
Корень как сеялка
Реферат
Корни растений питают почву водой и питательными веществами и преодолевают ее физическую компактность. Корни наделены механизмом, который позволяет им проникать и расти в плотных средах, таких как почва. Однако молекулярные механизмы, лежащие в основе этого процесса, все еще плохо изучены. Природа среды, в которой растут корни, усложняет анализ на месте механизмов, лежащих в основе проникновения корней.Подавление восприятия этилена путем внесения 1-метилциклопропена (1-MCP) на проростки томатов почти полностью исключило проникновение корней в Soilrite. Обращение этого процесса ауксином указывает на действие ауксин-этиленового сигнального пути в регуляции проникновения в корень. Мутант томата pct1–2 , который обнаруживает усиленный полярный транспорт ауксина, требует более высоких доз 1-MCP для ингибирования проникновения в корни, что указывает на ключевую роль транспорта ауксина в этом процессе.В этом обновлении мы представляем краткий обзор нашего текущего понимания молекулярных процессов, лежащих в основе проникновения корней у высших растений.
Ключевые слова: 1-MCP, транспорт ауксина, BSO, этилен, механический импеданс, pct1-2, PIN1, проникновение в корень, томат
Различия в привычках роста двух основных органов растения, корня и побега, в результате их принципиально разных ролей в приобретении ресурсов для завода. 1 Побег растет в воздухе практически без каких-либо препятствий, за исключением конкуренции с соседними растениями, в первую очередь за фиксацию углерода посредством фотосинтеза.Корень прорастает в почве для получения минеральных питательных веществ и воды, а также для прочного закрепления растения. 2 Почва, в которой корни растут и разрастаются, представляет собой плотную среду. Следовательно, корням требуется дополнительная способность расти в плотной среде, чтобы преодолевать физические ограничения, налагаемые структурой почвы. Плотная почва физически препятствует росту корней, а также снижает доступность воды и кислорода. 3 У сельскохозяйственных культур физические ограничения, налагаемые почвой, преодолеваются обработкой почвы перед посевом. Однако в природе семена прорастают и корни растут, преодолевая физические ограничения почвы. Корни демонстрируют высокую степень пластичности моделей роста, которые однозначно связаны с физическими и химическими свойствами почвы. Чтобы расти в подземной среде, корни наделены несколькими механизмами, которые регулируют и оптимизируют их рост. Одним из наиболее изученных механизмов является гравитропизм корней, 4 , вызывающий рост корней либо в направлении вектора гравитации, либо под углом, называемым плагиотропизмом.
Однако в природе семена прорастают и корни растут, преодолевая физические ограничения почвы. Корни демонстрируют высокую степень пластичности моделей роста, которые однозначно связаны с физическими и химическими свойствами почвы. Чтобы расти в подземной среде, корни наделены несколькими механизмами, которые регулируют и оптимизируют их рост. Одним из наиболее изученных механизмов является гравитропизм корней, 4 , вызывающий рост корней либо в направлении вектора гравитации, либо под углом, называемым плагиотропизмом.
По сути, считается, что в корнях гравитационный сигнал воспринимается через осаждение специализированных пластид, статолитов, в клетках колумеллы, расположенных в корневом покрове. 5 , 6 Физический сигнал осаждения статолита преобразуется в физиологическую информацию с помощью механизма, который еще предстоит полностью расшифровать, и передается в зону удлинения корня, расположенную на расстоянии от корня. Было идентифицировано несколько составляющих этого физиологического ответа, включая изменения pH, активных форм кислорода, индуцированную Ca +2 передачу сигналов и модуляцию транспорта ауксина. 7 — 9 Помимо силы тяжести, рост корней в почве регулируется несколькими другими факторами, позволяя корням определять наличие влаги и питательных веществ в почве и определять оптимальную стратегию роста. 19 , 20 Подобно направлению роста корня к вектору гравитации, восприятие сенсорных стимулов также играет важную роль в ориентации роста корня. 21 , 22 Кажется, что корни способны ощущать прикосновение и определять физическое сопротивление, оказываемое почвой на рост корней. 10 При встрече с препятствием, таким как твердый слой почвы, галька или другой корень, корни либо переориентируют свой рост, чтобы обогнуть препятствие (уклонение), либо развивают механическую силу, достаточную для проникновения через препятствие (преодоление). 23 В любом случае ощущение прикосновения является важным фактором, регулирующим рост корней. 24
23 В любом случае ощущение прикосновения является важным фактором, регулирующим рост корней. 24
Исследования биологии корней разных видов сельскохозяйственных культур показали, что даже в пределах одного и того же вида корни разных сортов различаются по своей способности проникать в более глубокие слои почвы для доступа к воде и питательным веществам. 11 — 13 Например, у сортов риса, способных проникать в твердыню, был лучший доступ к воде и они смогли избежать засухи при богарных землях, 14 , что указывает на то, что механическое сопротивление, испытываемое корнями, равно единице. основных ограничений роста корней в почве. 12 , 15 В соответствии с этим было обнаружено, что способность корней проникать в более прочные слои почвы контролируется генетически. 16 Несмотря на признание того, что реакция корней на механическое сопротивление имеет генетическую основу, однако, физиологический процесс, запускаемый в корнях для преодоления механического сопротивления, изучен плохо. 17 , 18
По сравнению с другими регулирующими факторами, имеется очень ограниченная информация о механизме, используемом корнями для определения механического импеданса, и связанной с ним передачи сигнала. Поскольку препятствия в почве первыми встречаются с клетками корневого чехлика, эти клетки являются основными кандидатами для восприятия стимулов прикосновения и / или механического сопротивления. 25 , 26 Хирургическое удаление кончика корня приводит к потере чувствительности и восстанавливается только после регенерации нового кончика корня. Клетки корневого чехлика находятся в динамическом состоянии потока, поскольку корневой покров постоянно сбрасывает эти клетки и заменяет их новыми, чтобы обеспечить беспрепятственный проход через почву. Эти клетки продуцируются корневой меристемой и дифференцируются в клетки, заполненные амилопластами, затем, наконец, становятся периферическими и в конечном итоге отслаиваются. 27 , 28 Этот транзит клеток через разные стадии развития привел к гипотезе о том, что относительное расположение этих клеток может определять их различную сенсорную активность. Данные показывают, что в то время как глубоко расположенные клетки колумеллы чувствуют силу тяжести, периферические клетки чувствуют прикосновение, о чем свидетельствует индуцированное прикосновением увеличение Ca +2 в этих клетках. 29 , 30 Однако эти две функции обнаружения могут перекрываться между двумя типами ячеек. 31 Несмотря на эти исследования, детали сигнальных систем, преодолевающих механический импеданс или вызванную прикосновением переориентацию в росте корня, остаются неуловимыми.
27 , 28 Этот транзит клеток через разные стадии развития привел к гипотезе о том, что относительное расположение этих клеток может определять их различную сенсорную активность. Данные показывают, что в то время как глубоко расположенные клетки колумеллы чувствуют силу тяжести, периферические клетки чувствуют прикосновение, о чем свидетельствует индуцированное прикосновением увеличение Ca +2 в этих клетках. 29 , 30 Однако эти две функции обнаружения могут перекрываться между двумя типами ячеек. 31 Несмотря на эти исследования, детали сигнальных систем, преодолевающих механический импеданс или вызванную прикосновением переориентацию в росте корня, остаются неуловимыми.
Учитывая потенциальное механистическое перекрытие между гравиперцепцией и механовосприятием, Тревавас и Найт 32 предположили, что грависенсинг представляет собой модифицированную систему сенсорного восприятия, поскольку оба ответа представляют собой своего рода механическое восприятие. В настоящее время считается, что рецептор прикосновения является механочувствительным каналом, и несколько линий доказательств указывают на существование активируемых растяжением каналов в плазматической мембране растений. 33 При восприятии сигнала прикосновения корни реагируют повышением цитозольного Ca 2+ , что, в свою очередь, вызывает временные изменения pH, внеклеточных ROS и изменения генома в паттернах экспрессии генов в тканях корня. 34 — 36 Считается, что повышение внеклеточного pH регулирует активность нескольких ферментов, влияющих на жесткость клеточной стенки. 37 , 38 Аналогичным образом, продукция ROS в апопласте может способствовать окислительному сшиванию компонентов клеточной стенки и, таким образом, укреплять клеточную стенку. 39 , 40 Ожидается, что эти модификации клеточной стенки у кончика корня могут придать корням необходимую жесткость для преодоления физических ограничений, обусловленных плотностью почвы.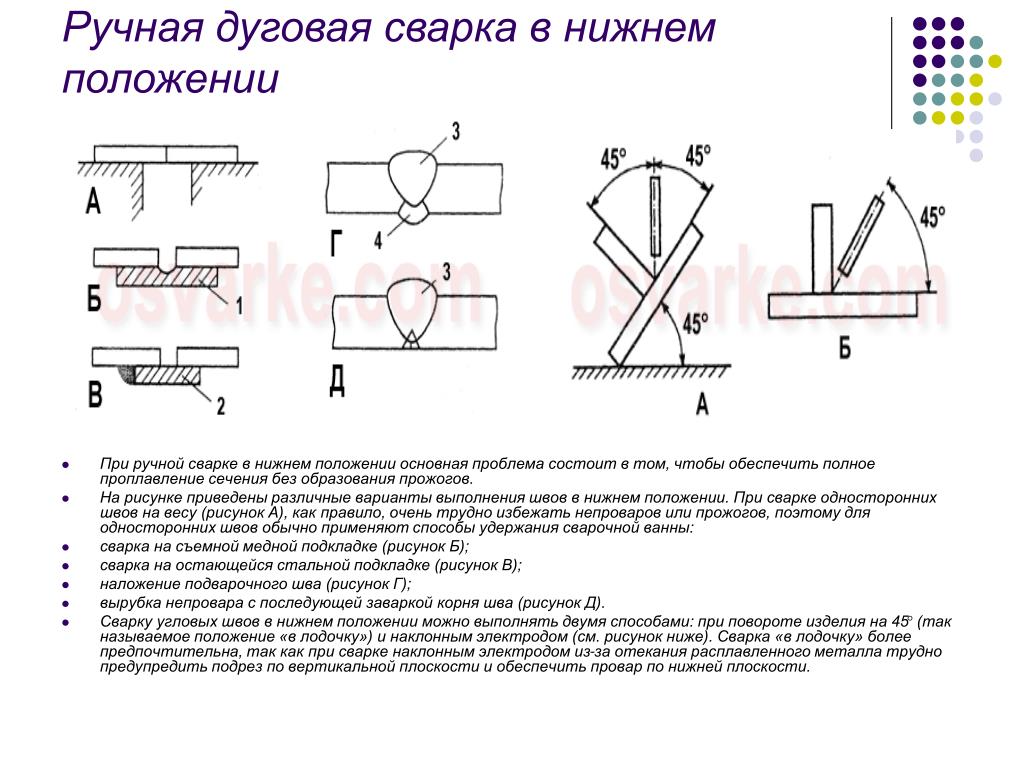
Хотя эти исследования подчеркивают регулирующую роль механовосприятия в росте корней, информации о меж- и внутриклеточной передаче сигналов в корнях и изменении морфологии корня при столкновении с препятствием мало. Одним из основных ограничений является сложность имитации почвенных условий при изучении этих реакций.В новаторском исследовании Окада и Шимура 41 провели скрининг мутантов роста корней Arabidopsis путем выращивания проростков на твердом агаре (1,5%) на чашках, наклоненных под углом 45 °. В то время как корни дикого типа демонстрировали волнообразный характер роста из-за реакции избегания препятствий, мутантные корни либо не обладали волнообразным характером роста, либо демонстрировали аномальный образец. Во многих случаях идентификация генов этих мутантов показывала дефекты транспорта ауксина, таким образом подчеркивая роль транспорта и передачи сигналов ауксина в преодолении механического сопротивления во время роста корня. 42 , 43
Okamoto et al. 44 исследовали рост и морфологию корней при механическом сопротивлении путем выращивания проростков арабидопсиса на горизонтальной поверхности чашки с агаром, покрытой диализной мембраной. Корни этих проростков не смогли проникнуть в субстрат и показали типичную морфологию, индуцированную этиленом, такую как уменьшение роста корней, связанное с уменьшением удлинения клеток и увеличением диаметра корня.Поскольку мутант eto1–1 с избыточным продуцированием этилена демонстрировал реакцию, аналогичную реакции дикого типа, а выработка этилена была аналогичной как в горизонтальных, так и в вертикальных условиях роста, похоже, что механический импеданс, вероятно, усиливал реакцию этилена в корнях, а не в корнях. биосинтез этилена. В соответствии с этой точкой зрения, этиленовые сигнальные мутанты ein2–1 и ctr1–1 не обнаруживают изменений в морфологии корня при воздействии механического импеданса.В соответствии с более ранними исследованиями Okada и Shimura, 42 Okamoto et al. 44 показали, что механический импеданс также изменяет реакцию на ауксин в корнях Arabidopsis, что визуализируется по более высокой экспрессии репортера DR5 :: GUS на нижней стороне кончика корня.
44 показали, что механический импеданс также изменяет реакцию на ауксин в корнях Arabidopsis, что визуализируется по более высокой экспрессии репортера DR5 :: GUS на нижней стороне кончика корня.
Взятые вместе, эти исследования согласуются с другими сообщениями, в которых было показано, что взаимодействие между передачей сигналов ауксина и этилена модулирует удлинение корня у Arabidopsis. Этилен регулирует несколько аспектов действия ауксина, например, положительную регуляцию акропетального и базипетального транспорта ауксина в корне, 45 — 47 и регулирует рост и развитие корня, контролируя биосинтез и транспорт ауксина в кончике корня. . 45 , 48 Такое синергетическое взаимодействие между ауксином и этиленом наблюдалось для нескольких специфических реакций, таких как регуляция гравитропизма корня, 49 рост корня, 50 развитие бокового корня 51 , 52 и дифференциация и удлинение корневых волосков. 53 Корни мутантов, таких как tir1 с дефектом восприятия ауксина, и pin2 и aux1 с дефектами оттока ауксина и переносчиков притока ауксина, соответственно, устойчивы к ингибированию роста в присутствии этилен. 54
Несмотря на наличие неопровержимых доказательств синергетического взаимодействия между ауксином и этиленом в регулировании развития и роста корней, в настоящее время мало что известно об относительной роли этих гормонов в регулировании проникновения корней в почву. 55 Текущие данные подтверждают, что этилен, по-видимому, играет важную роль в регулировании проникновения корней в почву. Роль этилена в проникновении в корни была продемонстрирована для томата, где корни проростков томатов, обработанных ингибиторами действия этилена, не могли проникать в 2% агар, но могли проникать в 0.5% агар. 56 В соответствии с этим, корень мутанта томата по восприятию этилена, Неспелые ( Nr ), демонстрирует пониженное проникновение в почву при наличии более высокого механического сопротивления. 57 Корни мутанта кукурузы Zmacs6 , дефектные по активности АСС-синтазы, фермента, ограничивающего скорость в пути биосинтеза этилена, также демонстрируют замедленный рост в почве, что свидетельствует о важности этилена в преодолении физического сопротивления. 58
57 Корни мутанта кукурузы Zmacs6 , дефектные по активности АСС-синтазы, фермента, ограничивающего скорость в пути биосинтеза этилена, также демонстрируют замедленный рост в почве, что свидетельствует о важности этилена в преодолении физического сопротивления. 58
В последние годы наличие специфических ингибиторов, которые блокируют гормональное восприятие или передачу сигналов, в значительной степени помогло в расшифровке сигнальных путей и идентификации молекул, регулирующих эти пути.Поскольку чрезмерное производство этилена во время хранения фруктов вызывает порчу таких фруктов, как банан, томат и яблоко, было разработано несколько специфических ингибиторов, блокирующих восприятие этилена. 59 Среди этих ингибиторов 1-метилциклопропан (1-MCP) является наиболее эффективным и, вероятно, блокирует действие этилена путем связывания с рецепторами этилена. 60 , 61
Корни проростков томатов, выращенных в присутствии 1-MCP, не смогли проникнуть в Soilrite.Поскольку 1-MCP также блокирует проникновение в корни однодольных растений, таких как рис и пшеница, и двудольных растений, таких как табак и салат, это указывает на роль этилена в регулировании проникновения в корни у широкого круга растений. 62 Ингибирующий эффект 1-MCP на проникновение в корень был обращен одновременным применением этилена, что подтверждает роль этиленовой передачи сигналов в регуляции проникновения в корень. Потеря проникновения в корень также была связана со снижением экспрессии репортера ауксинового ответа DR5 :: GUS в кончиках корней, что указывает на взаимодействие между передачей сигналов этилена и ауксина в регуляции этого ответа.Применение 1-MCP также снижает полярный транспорт ауксина в корнях томатов, предполагая, что взаимодействие ауксин-этилен может быть частично опосредовано модуляцией полярного транспорта ауксина. В соответствии с этой точкой зрения, мутант pct1-2 , который имеет почти в 3 раза более высокий транспорт ауксина, чем дикий тип, 63 , 64 показал близкое к нормальному проникновение корней в Soilrite в присутствии 1- МКП. Однако применение полярного ингибитора транспорта ауксина, TIBA, который препятствует перемещению везикул переносчиков ауксина, 65 вместе с 1-MCP частично ингибирует проникновение в корень мутанта pct1-2 .
Однако применение полярного ингибитора транспорта ауксина, TIBA, который препятствует перемещению везикул переносчиков ауксина, 65 вместе с 1-MCP частично ингибирует проникновение в корень мутанта pct1-2 .
Хотя обработка 1-MCP уменьшала транспорт ауксина в корнях pct1-2 , PAT оставалась значительно выше по сравнению с необработанными корнями дикого типа. Мы предположили, что увеличение дозировки 1-MCP будет постепенно блокировать PAT в корнях pct1-2 и будет препятствовать проникновению в корень даже в мутанте pct1-2 . В соответствии с нашим представлением, увеличение концентрации 1-MCP выше 4 мкл / л -1 ингибировало проникновение корней в pct1-2 до такой степени, что почти 50% корней не могли проникнуть в Soilrite ().Для сравнения, проникновение корней дикого типа в Soilrite почти полностью устраняется при 2 мкл л -1 . 62 В соответствии с наблюдениями, сделанными с диким типом, увеличение дозировки 1-MCP влияло также на рост гипокотилей и корней мутанта pct1-2 , стимулируя рост корней и ингибируя удлинение гипокотилей ().
Рис. 1. Дозовая реакция 1-MCP для pct1-2 проростков. pct1-2. проростков выращивали на Soilrite в присутствии и в отсутствие 1-MCP, как указано, при непрерывном белом свете в течение 7 дней.Проникновение в корни (A) и фенотип (B) проростков pct1-2 , выращенных в присутствии различных концентраций 1-MCP. Обратите внимание, что проростки pct1-2 демонстрируют частичное ингибирование проникновения корней, уменьшение длины гипокотилей и увеличение удлинения корня при концентрациях 1-MCP ≥ 4 мкл · л -1 . Звездочка (*) указывает на статистически значимую разницу в ответе на лечение (p <0,005 при n = 15–20 на группу).
Физиологические исследования показали, что система PAT требует активности специфических носителей притока и оттока ауксина, расположенных на плазматической мембране транспортирующих клеток. 66 , 67 , 68 Было показано, что у Arabidopsis и других растений белки PIN тесно связаны с полярным транспортом ауксина. 69 , 70 В Arabidopsis этилен регулирует транскрипцию нескольких транспортеров ауксина, включая PIN1, PIN2 и AUX1, 71, , 72 , таким образом, усиливая транспорт ауксина в корнях. Недавно мы продемонстрировали, что мутант pct1-2 демонстрирует повышенную экспрессию PIN1 в корне, что объясняет его повышенный полярный транспорт. 73 , 74 Для дальнейшего изучения связи между улучшенным PAT в pct1-2 и его устойчивостью к 1-MCP, мы обработали проростки pct1-2 L-бутионином- (S, R) -сульфоксимин (BSO), ингибитор биосинтеза глутатиона, который подавляет экспрессию нескольких белков PIN у арабидопсиса, таких как PIN1, PIN2 и PIN3. 75 , 76 Подобно заявке TIBA, применение 62 BSO вместе с 1-MCP вызывало снижение активности DR5 как в корнях дикого типа, так и в корнях pct1-2 ().Параллельно со снижением активности DR5 также подавлялось проникновение в корни, хотя гораздо более эффективно у дикого типа, чем у pct1-2 (). Эти результаты потенциально показывают, что снижение уровня ауксина и проникновение в корни 1-MCP связан с уменьшенной функцией PIN.
66 , 67 , 68 Было показано, что у Arabidopsis и других растений белки PIN тесно связаны с полярным транспортом ауксина. 69 , 70 В Arabidopsis этилен регулирует транскрипцию нескольких транспортеров ауксина, включая PIN1, PIN2 и AUX1, 71, , 72 , таким образом, усиливая транспорт ауксина в корнях. Недавно мы продемонстрировали, что мутант pct1-2 демонстрирует повышенную экспрессию PIN1 в корне, что объясняет его повышенный полярный транспорт. 73 , 74 Для дальнейшего изучения связи между улучшенным PAT в pct1-2 и его устойчивостью к 1-MCP, мы обработали проростки pct1-2 L-бутионином- (S, R) -сульфоксимин (BSO), ингибитор биосинтеза глутатиона, который подавляет экспрессию нескольких белков PIN у арабидопсиса, таких как PIN1, PIN2 и PIN3. 75 , 76 Подобно заявке TIBA, применение 62 BSO вместе с 1-MCP вызывало снижение активности DR5 как в корнях дикого типа, так и в корнях pct1-2 ().Параллельно со снижением активности DR5 также подавлялось проникновение в корни, хотя гораздо более эффективно у дикого типа, чем у pct1-2 (). Эти результаты потенциально показывают, что снижение уровня ауксина и проникновение в корни 1-MCP связан с уменьшенной функцией PIN.
Рис. 2. Количественная оценка влияния L-бутионин- (S, R) -сульфоксимина (BSO) на проникновение корней в проростках pct1-2 . Обратите внимание, что одновременное применение BSO вместе с 2 мкл L -1 1-MCP ингибирует экспрессию DR5 :: GUS в кончиках корней (A) и проникновение в корень (B) проростков pct1-2 .Проростки выращивали на Soilrite, смоченном 0,5 мМ BSO в присутствии или в отсутствие 1-MCP. Контрольные проростки выращивали на смоченном водой Soilrite. Звездочка (*) указывает на статистически значимую разницу в ответе на лечение (p <0,005 при n = 15–20 на группу).
Наши результаты указывают на тесное взаимодействие между этиленом и сигнальным путем ауксина в регуляции проникновения в корень. Наши результаты согласуются с сообщениями, показывающими, что у томатов мутант Nr с восприятием этилена показывает сниженный транспорт ИУК в корнях, а мутант epi , продуцирующий избыточный этилен, демонстрирует повышенный транспорт ИУК в корнях. 47 , 77 В этих сообщениях также предполагается, что этилен играет противоположную роль в транспорте ауксина в корнях и гипокотилях, с положительной регуляцией PAT в корнях и отрицательной модуляцией в ткани ствола. Точно так же мы также наблюдали некоторые контрастные ответы в корнях и гипокотилях томатов при обработке 1-МСР в отношении моделей роста и экспрессии генов, связанных с этиленом и ауксином. Тот факт, что применение 1-MCP уменьшало транспорт акропетального и базипетального ауксина в корнях томатов дикого типа, подтверждает, что этилен влияет на проникновение в корни и индуцированные проникновением изменения роста, по крайней мере частично, за счет увеличения полярного транспорта ауксина.Таким образом, наши результаты демонстрируют обязательное требование действия этилена для выполнения корневого проникновения в томате. Мы пришли к выводу, что этилен регулирует проникновение корней, по крайней мере частично, путем взаимодействия с ауксином. В будущем более глубокое знание реакции проникновения корней при импедансе должно помочь в разработке стратегий селекции сельскохозяйственных культур для улучшения проникновения корней и функции кормления для получения воды и питательных веществ для растений.
Проникновение корней томатов в почву требует взаимодействия между этиленом и ауксином.
- © 2011 Американское общество биологов растений
Реферат
Во время прорастания семян появляющиеся корни проявляют положительный гравитропизм и проникают в почву для питания и закрепления.Семена томата ( Solanum lycopersicum ), прорастающие в присутствии 1-метилциклопропена (1-MCP), ингибитора действия этилена, не смогли внедрить корни в Soilrite и росли на воздухе, образуя петли. Покадровая видеосъемка показала, что кончики корней, выращенные 1-MCP, сохраняют положительный гравитропизм и контактируют с поверхностью Soilrite, но не проникают в Soilrite. Исследования с течением времени показали, что эффект 1-MCP был наиболее заметным, когда набухание семян и проращивание проводились в постоянном присутствии 1-MCP.Напротив, 1-MCP был неэффективен при последующем проращивании после проникновения корней в Soilrite. Кроме того, обработка 1-MCP вызывала снижение активности ауксинового репортера DR5 :: β-глюкуронидазы и изменяла экспрессию транскриптов SlIAA3 и SlIAA9 , что указывает на вмешательство в передачу сигналов ауксина. Мутант с пониженным восприятием этилена, Never-ripe , продемонстрировал пониженную способность проникать в корни, а мутант с усиленным полярным транспортом ауксина, поликотиледон , показал почти нормальное проникновение в корни в присутствии 1-MCP, которое можно было обратить вспять с помощью применение ингибиторов транспорта ауксина.Наши результаты показывают, что во время прорастания семян томатов требуется совместное действие этилена и ауксина для проникновения корней в почву.
Покадровая видеосъемка показала, что кончики корней, выращенные 1-MCP, сохраняют положительный гравитропизм и контактируют с поверхностью Soilrite, но не проникают в Soilrite. Исследования с течением времени показали, что эффект 1-MCP был наиболее заметным, когда набухание семян и проращивание проводились в постоянном присутствии 1-MCP.Напротив, 1-MCP был неэффективен при последующем проращивании после проникновения корней в Soilrite. Кроме того, обработка 1-MCP вызывала снижение активности ауксинового репортера DR5 :: β-глюкуронидазы и изменяла экспрессию транскриптов SlIAA3 и SlIAA9 , что указывает на вмешательство в передачу сигналов ауксина. Мутант с пониженным восприятием этилена, Never-ripe , продемонстрировал пониженную способность проникать в корни, а мутант с усиленным полярным транспортом ауксина, поликотиледон , показал почти нормальное проникновение в корни в присутствии 1-MCP, которое можно было обратить вспять с помощью применение ингибиторов транспорта ауксина.Наши результаты показывают, что во время прорастания семян томатов требуется совместное действие этилена и ауксина для проникновения корней в почву.
Прорастание семян — решающий этап в жизненном цикле растений, когда проросшие проростки впервые сталкиваются с внешней средой и регулируют рост для оптимизации выживания. Как правило, первичный корень — это первый орган, который выходит из семени, и его последующий рост и развитие сильно зависят от местной окружающей среды. Корни растений выполняют многогранную роль, из которых две главные функции: прочно закрепить растение в почве и снабжать водой и питательными веществами надземные части растения (Hodge et al., 2009). Чтобы эффективно выполнять эти функции, корни наделены множеством сенсорных механизмов для определения силы тяжести, воды, питательных веществ и механического сопротивления почвы (Arnaud et al., 2010). Большое количество физиологических экспериментов в сочетании с хирургическим вскрытием показали, что кончик корня является местом восприятия и интеграции многих из этих сигналов (Iijima et al. , 2008; Arnaud et al., 2010). На кончике корня находится апикальная меристема корня, покоящийся центр и начальные клетки, которые дают начало всем типам клеток корня.Кончик корня окружен корневой крышкой, покрывающей апикальную меристему, которая состоит из двух тканей: колумеллы и клеток латерального корня крышки (Arnaud et al., 2010). Корневой покров обеспечивает физическую защиту апикальной меристемы корня от механического сопротивления при проникновении корней в почву. Кроме того, корневая крышка также функционирует как место для измерения силы тяжести, а хирургическое удаление и лазерная или генетическая абляция клеток корневой крышки приводят к потере гравитропного ответа (Blancaflor et al., 1998; Барлоу, 2003; Иидзима и др., 2003; Морита, 2010).
В соответствии с классической гипотезой Холодного-Вента, большое количество сообщений указывает на то, что дифференциальный рост корней после гравистимуляции инициируется асимметричным перераспределением ауксина растительного гормона (Rashotte et al., 2001; Ottenschläger et al., 2003 ; Мудай, Рахман, 2008; Ваннест, Фримл, 2009). Модель обратного фонтанного транспорта ауксина в верхушке корня предполагает, что ауксин, полученный из побегов, акропетально транспортируется в корневую крышку через сосудистую ткань корня, а после радиального распределения в корневой крышке в тканях эпидермиса он транспортируется обратно базипетально. зона удлинения корней, где он регулирует рост (Muday, De Long, 2001; Muday, Rahman, 2008; Vanneste, Friml, 2009).Исследования экспрессии репортеров ауксинового ответа, таких как DR5 :: GUS , показали, что максимальная активность DR5 в корне находится в области кончика корня, что указывает на более высокие уровни ауксина (Sabatini et al., 1999), который асимметрично распределяется во время гравитропного искривления корня (Rashotte et al., 2001; Ottenschläger et al., 2003). Рост удлинения корня также модулируется накоплением ауксина на кончике корня (Petersson et al., 2009), а накопление ауксина выше порогового уровня ингибирует удлинение корня. В соответствии с этим проростки мутанта с усиленным полярным транспортом ауксина томата ( Solanum lycopersicum ), поликотиледона ( pct1-2 ) имеют более короткие корни, а ингибирование транспорта ауксина у мутанта стимулирует удлинение корня (Al -Hammadi et al., 2003; Madishetty et al., 2006).
В соответствии с этим проростки мутанта с усиленным полярным транспортом ауксина томата ( Solanum lycopersicum ), поликотиледона ( pct1-2 ) имеют более короткие корни, а ингибирование транспорта ауксина у мутанта стимулирует удлинение корня (Al -Hammadi et al., 2003; Madishetty et al., 2006).
Хотя ауксин является наиболее важным гормоном, регулирующим рост корней, его действие модулируется рядом других гормонов, в частности, газообразным гормоном этиленом (Benková and Hejátko, 2009).Синергетическое взаимодействие между ауксином и этиленом было изучено в нескольких ответах развития корней у растений, таких как регуляция гравитропизма корней (Buer et al., 2006), рост корней (Rahman et al., 2001), развитие боковых корней (Ivanchenko et al., 2006). al., 2008, 2010; Negi et al., 2008), а также дифференциация и удлинение корневых волосков (Pitts et al., 1998). Один из способов взаимодействия — на уровне синтеза гормона, где ауксин стимулирует синтез этилена путем активации 1-аминоциклопропан-1-карбоксилат (АСС) синтазы, ключевого фермента в производстве этилена (Abel et al., 1995). Напротив, анализ корнеспецифических мутантов, нечувствительных к этилену, привел к открытию, что экспрессия генов, таких как ASA1 , ASB1 , TAA1 и TAA2 , которые участвуют в Trp-опосредованном биосинтезе ауксина, является активируется АЦЦ или этиленом в кончике корня (Степанова и др., 2005, 2008). Аналогичным образом, также сообщалось, что опосредованная этиленом повышающая регуляция переносчиков оттока ауксина, PIN1 и PIN2, и переносчика притока ауксина, AUX1, приводит к стимуляции базипетального транспорта ауксина к зоне удлинения корня у Arabidopsis ( Arabidopsis thaliana ) и томата (Rzicka et al., 2007; Степанова и др., 2007; Swarup et al., 2007; Неги и др., 2008, 2010). Этилен также ингибирует гравиответ корней, регулируя синтез флавоноидов, которые рассматриваются как вероятные регуляторы транспорта ауксина in vivo (Buer et al. , 2006). В кончике корня арабидопсиса этилен стимулирует биосинтез ауксина, что приводит к ингибированию удлинения корня (Růzicka et al., 2007; Swarup et al., 2007). Корни мутантов, таких как tir1 с дефектом восприятия ауксина, и pin2 и aux1 с дефектами транспортеров оттока ауксина и притока ауксина, соответственно, устойчивы к ингибированию роста в присутствии этилена ( Степанова и др., 2007). Сходное взаимодействие этилена-ауксина в регуляции транспорта и синтеза ауксина также описано для открытия крючка гипокотиля у Arabidopsis (Vandenbussche et al., 2010; Zádníková et al., 2010). Таким образом, хотя молекулярные детали, касающиеся способов взаимодействия между ауксиновым и этиленовым путями, все еще неясны, многочисленные исследования указывают на то, что существует обширная перекрестная связь между гормонами в регуляции роста корней.
, 2006). В кончике корня арабидопсиса этилен стимулирует биосинтез ауксина, что приводит к ингибированию удлинения корня (Růzicka et al., 2007; Swarup et al., 2007). Корни мутантов, таких как tir1 с дефектом восприятия ауксина, и pin2 и aux1 с дефектами транспортеров оттока ауксина и притока ауксина, соответственно, устойчивы к ингибированию роста в присутствии этилена ( Степанова и др., 2007). Сходное взаимодействие этилена-ауксина в регуляции транспорта и синтеза ауксина также описано для открытия крючка гипокотиля у Arabidopsis (Vandenbussche et al., 2010; Zádníková et al., 2010). Таким образом, хотя молекулярные детали, касающиеся способов взаимодействия между ауксиновым и этиленовым путями, все еще неясны, многочисленные исследования указывают на то, что существует обширная перекрестная связь между гормонами в регуляции роста корней.
Несмотря на важную роль ауксина и этилена в регуляции развития и роста корней, имеется мало информации об относительных функциях этих гормонов в регулировании проникновения корней в почву (Clark et al., 2003; Бенкова и Хеджатко, 2009; Ходж и др., 2009). Исследование корней, подвергшихся механическому сопротивлению проникновению в среду, показало, что увеличение синтеза этилена и / или передачи сигналов опосредует уменьшение роста корня, что сопровождается увеличением диаметра корня и уменьшением удлинения клеток корня (Sarquis et al. , 1991; Okamoto et al., 2008). Эти морфологические изменения, вероятно, увеличивают способность корней преодолевать физическое сопротивление при проникновении в почву. Как показано на примере корней кукурузы ( Zea mays ), корневая оболочка играет важную роль в этом процессе, поскольку она снижает трение и способствует проникновению корней в почву (Iijima et al., 2003). Было показано, что локальный синтез ауксина и этилена происходит в кончике корня. Например, транскрипты АСС-синтазы и АСС-оксидазы, ключевых ферментов, регулирующих биосинтез этилена, были обнаружены в кончиках корней кукурузы (Gallie et al. , 2009). Сходным образом ауксин синтезируется в верхушке корня Arabidopsis (Růzicka et al., 2007; Swarup et al., 2007). Ингибирование действия этилена усиливает рост корней кукурузы, указывая на то, что эндогенно продуцируемый этилен вызывает уменьшение роста корней (Whalen and Feldman, 1988).Экзогенно применяемый этилен быстро подавляет рост корней, ингибируя удлинение клеток в области, проксимальной к кончику корня у кукурузы (Whalen and Feldman, 1988) и Arabidopsis (Le et al., 2001). Корни мутанта кукурузы Zmacs6 , дефектного по активности АСС-синтазы, демонстрируют замедленный рост в почве, что свидетельствует о важности этилена в преодолении физической устойчивости (Gallie et al., 2009). Корни проростков томатов, обработанные ингибиторами действия этилена, также не могут проникать в 2% агар, но могут проникать 0.5% агар (Zacarias and Reid, 1992). Точно так же нечувствительный к этилену мутант томата Неспелые ( Nr ) демонстрирует пониженную способность проникать в почву при наличии более высокого механического сопротивления (Clark et al., 1999). Генетические подходы применялись для идентификации локусов количественных признаков (QTL), ответственных за проникновение корней в почву, особенно у таких сельскохозяйственных культур, как рис ( Oryza sativa ; Price et al., 2000). Куртуа и др. (2009) объединили исследования по обнаружению QTL, проведенные разными группами у риса по различным корневым признакам, с использованием мета-QTL-анализа, охватывающего весь геном, и идентифицировали 35 QTL для индекса проникновения в корень у риса.Ожидается, что в ближайшее время будут идентифицированы гены, регулирующие проникновение корней у сельскохозяйственных культур.
, 2009). Сходным образом ауксин синтезируется в верхушке корня Arabidopsis (Růzicka et al., 2007; Swarup et al., 2007). Ингибирование действия этилена усиливает рост корней кукурузы, указывая на то, что эндогенно продуцируемый этилен вызывает уменьшение роста корней (Whalen and Feldman, 1988).Экзогенно применяемый этилен быстро подавляет рост корней, ингибируя удлинение клеток в области, проксимальной к кончику корня у кукурузы (Whalen and Feldman, 1988) и Arabidopsis (Le et al., 2001). Корни мутанта кукурузы Zmacs6 , дефектного по активности АСС-синтазы, демонстрируют замедленный рост в почве, что свидетельствует о важности этилена в преодолении физической устойчивости (Gallie et al., 2009). Корни проростков томатов, обработанные ингибиторами действия этилена, также не могут проникать в 2% агар, но могут проникать 0.5% агар (Zacarias and Reid, 1992). Точно так же нечувствительный к этилену мутант томата Неспелые ( Nr ) демонстрирует пониженную способность проникать в почву при наличии более высокого механического сопротивления (Clark et al., 1999). Генетические подходы применялись для идентификации локусов количественных признаков (QTL), ответственных за проникновение корней в почву, особенно у таких сельскохозяйственных культур, как рис ( Oryza sativa ; Price et al., 2000). Куртуа и др. (2009) объединили исследования по обнаружению QTL, проведенные разными группами у риса по различным корневым признакам, с использованием мета-QTL-анализа, охватывающего весь геном, и идентифицировали 35 QTL для индекса проникновения в корень у риса.Ожидается, что в ближайшее время будут идентифицированы гены, регулирующие проникновение корней у сельскохозяйственных культур.
Чтобы лучше понять относительный вклад этилена и ауксина в процесс проникновения в корни, мы исследовали этот ответ у томатов дикого типа и у мутанта с усиленным полярным транспортом ауксина, pct1-2 , в условиях пониженной передачи сигналов этилена. Мы также проанализировали взаимодействие между этими двумя гормонами, используя ауксин-чувствительный репортерный ген и изучив экспрессию этиленовых и сигнальных генов ауксина с помощью количественной обратной транскрипции (qRT) -PCR. Результаты, полученные в этом исследовании, демонстрируют, что совместное действие ауксина и этилена необходимо для проникновения корней томатов в почву.
Результаты, полученные в этом исследовании, демонстрируют, что совместное действие ауксина и этилена необходимо для проникновения корней томатов в почву.
РЕЗУЛЬТАТЫ
Ингибирование сигналов этилена ухудшает проникновение корней в почву
Чтобы изучить роль этилена в росте корней томатов, мы проращивали семена томатов на Soilrite в присутствии 1-метилциклопропена (1-MCP), ингибитора рецепторы этилена (Sisler et al., 1996; Sisler, 2006). Удивительно, но в отличие от контрольных сеянцев, которые показали нормальное проникновение, корни проростков, обработанных 1-MCP, не смогли проникнуть в Soilrite и росли на воздухе, образуя петли (рис.1А). Чтобы убедиться, что наблюдаемое отсутствие проникновения в корни в присутствии 1-MCP было конкретно связано с ингибированием рецепторов этилена, мы исследовали проникновение в корни у мутанта томата Nr , который несет усеченную версию в одном из рецепторов этилена томата. , ETR3 (Wilkinson et al., 1995). Большинство необработанных проростков Nr (67%) показали недостаточное проникновение корней на Soilrite, тогда как проростки Nr , обработанные 1-MCP, показали полную потерю проникновения корней (рис.1B; Дополнительный рис. S1A). Уменьшение проникновения корня в Nr на Soilrite согласуется с более ранним исследованием Clark et al. (1999), которые сообщили о потере 47% проникновения корней в Nr на песчаной среде. В целом эти наблюдения показывают, что 1-MCP ингибирует проникновение в корни из-за ингибирования регулируемых этиленом путей.
Рисунок 1. Ингибирование передачи сигналов этилена препятствует проникновению корней томатов. A. Фенотипы проростков томатов, выращенных в присутствии или в отсутствие 1-MCP в течение 5 дней.Обратите внимание, что корни проростков, обработанных 1-MCP, не попали в Soilrite и росли в воздухе (стрелка). B, Отсутствие прорастания корней и воздушного роста корней (стрелки) у проростков Nr . Обратите внимание на преувеличенный ответ на лечение 1-MCP. C, выделение этилена (Et) из проростков томатов ( n = 15–20), выращенных в присутствии или в отсутствие 1-MCP. Через указанные временные интервалы собирали 1 мл газа, свободного от паровой фазы, и определяли концентрацию этилена. D. Одновременное применение этилена с 1-MCP блокирует ингибирующий эффект 1-MCP дозозависимым образом.Проникновение корней контролировали у семидневных проростков томатов ( n = 20). Звездочки указывают на статистически значимые различия между контрольными проростками и проростками, обработанными 1-MCP (однофакторный дисперсионный анализ; P <0,005). [Цветную версию этого рисунка см. В статье в Интернете.]
C, выделение этилена (Et) из проростков томатов ( n = 15–20), выращенных в присутствии или в отсутствие 1-MCP. Через указанные временные интервалы собирали 1 мл газа, свободного от паровой фазы, и определяли концентрацию этилена. D. Одновременное применение этилена с 1-MCP блокирует ингибирующий эффект 1-MCP дозозависимым образом.Проникновение корней контролировали у семидневных проростков томатов ( n = 20). Звездочки указывают на статистически значимые различия между контрольными проростками и проростками, обработанными 1-MCP (однофакторный дисперсионный анализ; P <0,005). [Цветную версию этого рисунка см. В статье в Интернете.]
Учитывая естественную склонность зарождающихся корней врастать в почву, было интересно изучить, может ли 1-MCP вызывать рост корней в воздухе у других видов, кроме томатов. Корни однодольных проростков пшеницы ( Triticum aestivum ; дополнительный рис.S1B) и риса (данные не показаны), а также проростки двудольных растений, табака ( Nicotiana tabacum ) и салата ( Lactuca sativa ; данные не показаны), выращенные в присутствии 1-MCP, также показали аналогичный рост корней в воздухе с формирование петли. У пшеницы и риса при прорастании появлялось от трех до шести осей первичных корней, и в присутствии 1-MCP все образовывали воздушные петли. В отсутствие 1-MCP корни соответствующих контрольных проростков проникали нормально и росли внутри Soilrite.
Исследование выделения этилена из проростков показало, что 1-MCP не влиял на этот процесс до стадии прорастания корешка, то есть примерно через 48 часов после посева.Однако после появления корешка через 3 дня после посева выделение этилена из проростков, обработанных 1-MCP, было снижено по сравнению с контрольными проростками (рис. 1С). Мы также исследовали, может ли экзогенное применение этилена обратить вспять воздушный рост корней, вызванный 1-MCP, и позволить корням проникнуть в Soilrite. На рисунке 1D показано, что проростки, прорастающие в присутствии как 1-MCP, так и этилена, демонстрируют увеличение проникновения корней с увеличением концентрации этилена, что указывает на то, что этилен может противодействовать индуцированному 1-MCP ингибированию проникновения в корни. Это согласуется с предыдущими сообщениями о том, что экзогенно применяемый этилен может частично восстанавливать индуцированное 1-МСР ингибирование рецепторов этилена у некоторых видов растений (Sisler et al., 1996; Binder and Bleecker, 2003; Feng et al., 2004; Sisler, 2006). ). Взятые вместе, эти наблюдения убедительно подтверждают роль этилена в регулировании проникновения корней.
Это согласуется с предыдущими сообщениями о том, что экзогенно применяемый этилен может частично восстанавливать индуцированное 1-МСР ингибирование рецепторов этилена у некоторых видов растений (Sisler et al., 1996; Binder and Bleecker, 2003; Feng et al., 2004; Sisler, 2006). ). Взятые вместе, эти наблюдения убедительно подтверждают роль этилена в регулировании проникновения корней.
1-МСР вызывает специфические реакции роста, связанные с передачей сигналов этилена
Известно, что воздействие этилена приводит к стимулированию удлинения гипокотилей у выращенных на свету проростков (Smalle et al., 1997; Vandenbussche et al., 2003a, 2003b), уменьшение гравитропической кривизны (Buer et al., 2006) и ингибирование удлинения корня у нескольких видов (Smalle and van der Straeten, 1997; Buer et al., 2006). Чтобы еще раз подтвердить, что 1-MCP действует в томатах, ингибируя действие этилена, мы исследовали влияние различных концентраций 1-MCP на удлинение гипокотилей и корней проростков томатов, выращенных на Soilrite. На рис. 2А показано, что 1-МСР вызывает прогрессирующее дозозависимое уменьшение длины гипокотиля при одновременном увеличении удлинения корней.Одновременное воздействие этилена частично сводило на нет вышеуказанные реакции роста 1-MCP (дополнительный рисунок S1C). Влияние 1-MCP на ростовую реакцию также наблюдалось у проростков, выращенных на вертикальных чашках с агаром (дополнительный рис. S1D), демонстрируя, что эти эффекты не возникали вторично из-за нарушения проникновения корней. Корни проростков, обработанных 1-MCP, а также контрольных проростков, выращенных на вертикальных чашках с агаром, были более вытянутыми, чем у проростков, выращенных на Soilrite, вероятно, из-за отсутствия какого-либо физического сопротивления росту корней, а также из-за различий в условиях роста. .Эффект 1-MCP на рост проростков был наиболее заметным, когда наблюдалось изменение отношения длины корня к длине гипокотиля. В среднем соотношение длины корня к длине гипокотиля 2,84 и 4,4 наблюдали для необработанных проростков, выращенных на пластинах Soilrite или вертикальных агаровых чашках, соответственно, и гораздо более высокие отношения 9,87 и 10,6 наблюдались при добавлении 1-MCP ( P <0,005; n = 15). Таким образом, 1-MCP действует в томатах противоположным образом в гипокотилях и корнях, что известно о влиянии этилена на различные виды растений.
Таким образом, 1-MCP действует в томатах противоположным образом в гипокотилях и корнях, что известно о влиянии этилена на различные виды растений.
1-MCP оказывает специфическое влияние на рост и развитие сеянцев томатов. A. Влияние различных концентраций 1-MCP на длину корня и гипокотиля (Hypo) семидневных проростков томатов. B. Сравнение действия 1-MCP (2 мкл L –1 ) на длину корня и гипокотиля 7-дневных проростков томатов дикого типа (WT) и Nr . C. Изображения 9-дневных проростков томатов, выращенных на вертикальных пластинах с агаром в присутствии или в отсутствие 1-MCP, демонстрируют усиленное развитие корней и продвинутый органогенез первичных листьев (стрелка).D. Количественная оценка влияния 1-MCP на формирование боковых корней у проростков, выращенных на вертикальных пластинах с агаром. E, отсутствие эффекта 1-MCP на прорастание семян томатов. Семена высевали на агар в присутствии или в отсутствие 1-MCP, и за прорастанием следили путем количественной оценки появления корешка из семенной оболочки в моменты времени, указанные на оси абсцисс. F. Эффект 1-MCP на проникновение корня в Soilrite зависит от времени применения. В моменты времени, указанные на оси абсцисс, добавляли 1-MCP и коробки закрывали.Процент проникновения корней определяли через 5 дней после посева. Звездочки и решетки указывают на статистически значимые различия между обработанными и контрольными растениями, а также между растениями дикого типа и мутантными растениями, соответственно ( P <0,005; n = 15–30 на группу). [См. Онлайн-статью для цветной версии этого рисунка.]
Чтобы дополнительно проверить, связаны ли эффекты развития, вызванные 1-MCP, с передачей сигналов этилена, мы исследовали рост и ответы 1-MCP мутанта Nr .На контрольной среде корни Nr были длиннее, чем корни дикого типа, и воздействие 1-MCP не увеличивало, а скорее немного уменьшало длину корня Nr (фиг. 2B). Интересно, что гипокотили Nr имели сходную длину по сравнению с диким типом и демонстрировали нормальное ингибирование удлинения в присутствии 1-МСР. Это может быть связано с различным относительным участием рецепторов ETR в регуляции удлинения корня и гипокотиля. Хотя ожидается, что 1-MCP воздействует на несколько рецепторов этилена, в Nr мутирован только рецептор ETR3, хотя нефункциональный рецептор может ингибировать другие рецепторы этилена, связываясь с ними (Hackett et al., 2000).
Это может быть связано с различным относительным участием рецепторов ETR в регуляции удлинения корня и гипокотиля. Хотя ожидается, что 1-MCP воздействует на несколько рецепторов этилена, в Nr мутирован только рецептор ETR3, хотя нефункциональный рецептор может ингибировать другие рецепторы этилена, связываясь с ними (Hackett et al., 2000).
Действие 1-MCP на проростки томатов не ограничивалось модификацией роста корней и гипокотилей; лечение также стимулировало органогенез. Проростки, обработанные 1-MCP, инициировали большее количество боковых корней (рис. 2, C и D), проявляя фенотип, аналогичный фенотипу, описанному для Nr (Negi et al., 2010), и показали более быстрое появление первичных листьев на апикальная меристема побега (рис. 2В). Однако 1-MCP не влиял на зарождение придаточных корней, что было аналогично таковому у контрольных проростков (данные не показаны).Точно так же 1-MCP не влиял на время прорастания семян, которое было сходным в обработанных и необработанных образцах (рис. 2E). Эти наблюдения демонстрируют, что 1-MCP влияет на определенные аспекты роста и развития проростков томатов и действует в томатах, ингибируя этиленовые реакции.
Эффект 1-MCP наиболее силен при применении до начала прорастания корней
После посева семян томатов обычно требуется не менее 24 часов для прорастания корешка из семян.Перед появлением корешок д. Преодолевать механический барьер из четырех слоев клеток в крышке эндосперма и слое семенников (Toorop et al., 2000). Чтобы точно определить стадию развития, на которой 1-MCP запускал максимальное ингибирование проникновения корней в Soilrite, семена томатов переносили на 1-MCP либо сразу после посева, либо через прогрессивные интервалы времени после посева. На рис. 2F показано, что наибольшее ингибирование проникновения корней наблюдалось, когда 1-MCP применялся непосредственно после посева или до проникновения корней в Soilrite.Ингибирующий эффект 1-MCP был резко снижен, когда он применялся после проникновения корня. В таких случаях корни продолжали расти в Soilrite и не выходили из Soilrite (данные не показаны). По-видимому, 1-MCP был наиболее эффективным в блокировании проникновения корней, когда присутствовал до начальной стадии проникновения корней в Soilrite.
В таких случаях корни продолжали расти в Soilrite и не выходили из Soilrite (данные не показаны). По-видимому, 1-MCP был наиболее эффективным в блокировании проникновения корней, когда присутствовал до начальной стадии проникновения корней в Soilrite.
Корни, обработанные 1-MCP, сохраняют нормальную гравитропную реакцию
Корни имеют положительную гравитропную природу, и эта реакция необходима для роста корней в почву.Мутанты Arabidopsis, такие как aux1 , которые дефектны по гравитропизму корней, демонстрируют корни, которые беспорядочно растут на поверхности среды (Bennett et al., 1996). Поэтому мы рассмотрели возможность того, что корни проростков, проросших в присутствии 1-MCP, росли на воздухе, образуя петли, поскольку они утратили свой гравитропный отклик. Используя покадровую визуализацию, мы сравнили динамику гравитропической кривизны корней необработанных и обработанных 1-MCP проростков томатов, выращенных на вертикальных пластинах с агаром, после ориентации проростков по горизонтали.Мы не обнаружили существенной разницы в гравитропной кривизне корня между обработанными и необработанными сеянцами (фиг. 3A), что указывает на то, что 1-MCP не оказывал никакого влияния на гравитропный отклик корня при испытанных концентрациях.
Рисунок 3. 1-MCP ингибирует проникновение корней в проростках томатов независимо от какого-либо воздействия на гравитропный или фототропный ответ. A. Гравитропный ответ в корнях проростков, выращенных в присутствии или в отсутствие 1-MCP. Проростки, выращенные на вертикальных пластинах с агаром, переориентировали под углом 90 ° относительно вектора силы тяжести.Кривизну кончика корня определяли путем фотографирования корней через заданные промежутки времени с помощью камеры. B, покадровая съемка проростков томатов, выращенных на Soilrite, в присутствии и в отсутствие 1-MCP. Изображения были воспроизведены из дополнительного фильма S1 (-MCP) и дополнительного фильма S2 (+ MCP). Проростки выращивали в плотно закрытых ящиках при однонаправленном белом свете в течение 4 дней. Кадры были захвачены в указанные моменты времени. У контрольных проростков (–МЦП) отсутствует воздушный рост корней, стрелки указывают на гипокотили.У обработанных проростков (+ MCP) стрелки указывают на один и тот же репрезентативный корень в разные моменты времени. Обратите внимание, что кончик корня является положительно гравитропным и все время остается в тесном контакте с поверхностью Soilrite. Дополнительные сведения см. В разделе «Дополнительные фильмы S1 и S2». [Цветную версию этого рисунка см. В статье в Интернете.]
Кадры были захвачены в указанные моменты времени. У контрольных проростков (–МЦП) отсутствует воздушный рост корней, стрелки указывают на гипокотили.У обработанных проростков (+ MCP) стрелки указывают на один и тот же репрезентативный корень в разные моменты времени. Обратите внимание, что кончик корня является положительно гравитропным и все время остается в тесном контакте с поверхностью Soilrite. Дополнительные сведения см. В разделе «Дополнительные фильмы S1 и S2». [Цветную версию этого рисунка см. В статье в Интернете.]
Чтобы иметь возможность отслеживать гравитропный отклик корня одновременно с проникновением корня, мы выполнили покадровую визуализацию проростков томатов, выращенных на Soilrite в присутствии или в отсутствие 1-MCP ( Дополнительные фильмы S1 и S2).Было очевидно, что, хотя корни проростков, обработанных 1-MCP, не могли проникнуть в Soilrite и образовали воздушные петли, кончики корней все время оставались в тесном контакте с Soilrite, явно демонстрируя положительный гравитропизм (Рис. 3B; Дополнительный фильм S2). По мере того, как корни удлинялись, они отображали гораздо более крупные петли, но их кончики все еще оставались в тесном контакте с поверхностью Soilrite. Такое поведение корня аналогично тому, что наблюдается у Arabidopsis, когда при столкновении с препятствием корень образует изгиб, позволяя корневому покрову оставаться в контакте с объектом, в то время как главный корень удлиняется параллельно поверхности препятствия (Масса и Гилрой , 2003).Таким образом, в проростках, выращенных на Soilrite, 1-MCP ингибировал проникновение корней, но не оказывал ингибирующего действия на гравитропный ответ корня при тестируемых концентрациях.
Используя однонаправленный источник света во время покадровой съемки, мы также смогли проанализировать влияние 1-MCP на фототропную реакцию гипокотиля во время процесса проникновения корня. Мы не обнаружили какой-либо существенной разницы между сеянцами, обработанными 1-MCP, и необработанными (рис. 3B; дополнительные видеоролики S1 и S2).И обработанные, и необработанные проростки показали ориентацию гипокотилей в направлении света, показывая нормальные фототропные реакции. Хотя конденсация влаги в запечатанных коробках размывает изображение на 4 дня, фенотипы отчетливо видны. Взятые вместе, эти наблюдения показывают, что 1-MCP может препятствовать проникновению корней через процесс, который не зависит от воздействия на гравитропные или фототропные реакции.
Лечение 1-MCP вызывает изменения в экспрессии генов, связанных с этиленом, ауксином и механоцепцией
Мы спросили, какие сигнальные компоненты участвуют в способности 1-MCP ингибировать проникновение в корень.Способность этилена способствовать удлинению гипокотилей у выращенных на свету растений зависит от передачи сигналов ауксина (Vandenbussche et al., 2003a, 2003b). Точно так же ингибирование этиленом удлинения корней у этиолированных проростков коррелирует с экспрессией генов, связанных как с этиленом, так и с ауксином (Степанова, Алонсо, 2005; Степанова и др., 2007). Предполагается, что путь передачи сигнала, регулирующий ингибирование роста корней этиленом, включает линейную последовательность событий с использованием сначала этиленовых, а затем ауксиновых сигнальных компонентов (Roman et al., 1995). Поэтому мы сначала исследовали, влияет ли 1-MCP на рецепторы этилена томатов на уровне транскрипции в дополнение к его известному влиянию на активность рецепторов. Транскрипты всех шести рецепторов ETR были обнаружены в корнях томатов с помощью qRT-PCR, а четыре рецептора, ETR2 , -4 , -5 и — 6 , показали заметное повышение экспрессии в корнях после 1 -MCP, тогда как экспрессия ETR1 и NR ( ETR3 ) была снижена.Для сравнения, экспрессия NR , ETR4 , -5 и — 6 была снижена, а экспрессия ETR1 и ETR2 была повышена в гипокотилях, обработанных 1-MCP ( Рис.4, А и Б). По-видимому, воздействие 1-MCP влияло на экспрессию ETR как в гипокотилях, так и в корнях, хотя эффект 1-MCP варьировал в разных тканях, в некоторой степени аналогично его дифференциальному эффекту на удлинение корня и гипокотиля, обсуждавшемуся выше (рис. 2B).
По-видимому, воздействие 1-MCP влияло на экспрессию ETR как в гипокотилях, так и в корнях, хотя эффект 1-MCP варьировал в разных тканях, в некоторой степени аналогично его дифференциальному эффекту на удлинение корня и гипокотиля, обсуждавшемуся выше (рис. 2B).
1-MCP влияет на экспрессию генов, связанных с этиленом, ауксином и механоцепцией, в проростках томатов. A и B, Количественная оценка относительных уровней экспрессии генов в кончиках корней (A) и гипокотилях (B). РНК для анализа экспрессии генов выделяли из трехдневных проростков, выращенных в присутствии или в отсутствие 1-МСР. Уровни экспрессии членов семейств генов ETR и IAA и гена, подобного MCA1 , количественно оценивали с помощью qRT-PCR. Относительный уровень экспрессии на оси y относится к кратной разнице в экспрессии каждого гена по сравнению с необработанным контролем, взятым в качестве эталона.Данные соответствуют средним значениям трех повторов ± стандартная ошибка. C и D, Экспрессия репортера ауксина DR5 :: GUS в корнях, выращенных в присутствии или в отсутствие 1-MCP. Кончики корней анализировали через 3 дня (C) и 5 дней (D) после посева. [См. Онлайн-статью для цветной версии этого рисунка.]
У томатов информация о взаимодействии между сигнальными путями ауксина и этилена ограничена. Однако анализ экспрессии показал, что два сигнальных гена ауксина, SlIAA3 и SlIAA9 , играют важную роль в опосредованной этиленом регуляции развития плода и открытия крючков гипокотиля, соответственно (Wang et al., 2005; Chaabouni et al., 2009). Изучая уровни транскриптов этих двух генов, мы обнаружили, что их экспрессия была увеличена в кончиках корней проростков, обработанных 1-MCP. Напротив, в гипокотилях уровень SlIAA3 был повышен, а уровень транскрипта SlIAA9 был снижен обработкой 1-MCP, демонстрируя дифференциальные ответы, как наблюдали с транскриптами ETR .
Мы также спросили, может ли 1-MCP ингибировать проникновение в корни, подавляя экспрессию генов, участвующих в механовосприятия.Хотя до настоящего времени не было идентифицировано специфических механосенсоров растений, имеются доказательства, что активируемые прикосновением ионные каналы в плазматической мембране участвуют в реакции растений на прикосновение (Monshausen et al., 2008). У Arabidopsis проникновение кончиков корней в среду было тесно связано с экспрессией гена MID1-COMPLEMENTING ACTIVITY1 ( MCA1 ), механочувствительного канала, проницаемого для Ca 2+ (Nakagawa et al., 2007). Используя транскрипт MCA1 Arabidopsis для выравнивания последовательностей, мы идентифицировали предполагаемый транскрипт MCA1 томата (EST SGN-E 1245095 в Unigene SGN-U565830) с высокой гомологией с AtMCA1 .qRT-PCR показала, что экспрессия гена MCA1 -подобного томата была повышена в обработанных 1-MCP кончиках корней (рис. 4A), тогда как в гипокотилях его экспрессия снижалась при обработке 1-MCP (рис. 4B). ). Поскольку MCA1 -подобный транскрипт томата активируется в корнях, обработанных 1-MCP, маловероятно, что неспособность корней проникать в среду была вызвана ингибированием механовосприятия. В целом, эти данные показывают, что 1-MCP модулирует экспрессию генов, связанных с этиленом, ауксином и механоцепцией, одновременно с ингибированием проникновения в корень.
Лечение 1-MCP, вероятно, изменяет накопление ауксина в кончике корня
Учитывая наблюдаемые изменения в экспрессии транскриптов гена ауксинового ответа после лечения 1-MCP, необходимо было более подробно изучить корреляцию между ингибированием этилена. передача сигналов 1-MCP, проникновение в корень и изменение ответа на ауксин в кончике корня. Несколько исследований показали, что активность синтетического консенсусного репортера ауксина, DR5 :: GUS , точно отражает индуцированную ауксином экспрессию гена в тканях (Ulmasov et al. , 1997; Sabatini et al., 1999). Анализируя корни, выросшие в присутствии или в отсутствие 1-MCP, мы обнаружили, что репортер DR5 четко продемонстрировал разницу между обработанными и контрольными сеянцами. В корнях контрольных проростков максимальная активность DR5 наблюдалась в кончиках корней, и активность постепенно снижалась до неопределяемых уровней по направлению к зоне удлинения. В проростках, обработанных 1-MCP, активность DR5 была менее интенсивной и распространялась на гораздо меньшую область. Подобные различия наблюдались как у 3-, так и у 5-дневных проростков (рис.4, В и Г). Эти данные продемонстрировали, что активность DR5 может быть использована для исследования изменений в реакции на ауксин во время индуцированного 1-MCP ингибирования проникновения корней в Soilrite.
, 1997; Sabatini et al., 1999). Анализируя корни, выросшие в присутствии или в отсутствие 1-MCP, мы обнаружили, что репортер DR5 четко продемонстрировал разницу между обработанными и контрольными сеянцами. В корнях контрольных проростков максимальная активность DR5 наблюдалась в кончиках корней, и активность постепенно снижалась до неопределяемых уровней по направлению к зоне удлинения. В проростках, обработанных 1-MCP, активность DR5 была менее интенсивной и распространялась на гораздо меньшую область. Подобные различия наблюдались как у 3-, так и у 5-дневных проростков (рис.4, В и Г). Эти данные продемонстрировали, что активность DR5 может быть использована для исследования изменений в реакции на ауксин во время индуцированного 1-MCP ингибирования проникновения корней в Soilrite.
Затем мы исследовали во временном разрезе, восстановит ли удаление 1-MCP уровень экспрессии DR5 в кончиках корней томатов вместе с возобновлением проникновения корней в Soilrite. Для этого мы сначала выращивали проростки в течение 3 дней в присутствии 1-MCP, а затем выращивали в открытых ящиках при нормальных атмосферных условиях.Через 12 часов после удаления 1-MCP корни начали проникать в Soilrite, и почти все корни проникли в течение 48 часов (рис. 5A). Одновременно наблюдалось увеличение активности DR5 в кончиках корней с максимумом через 24 часа (рис. 5B), момент времени, когда почти 60% корней вошли в Soilrite (рис. 5A). После этого, хотя активность DR5 снизилась аналогичным образом как у обработанных, так и у необработанных проростков, корни продолжали проникать в Soilrite, и процесс был почти завершен к 48 часам.Взятые вместе, эти наблюдения предполагают вероятное взаимодействие между передачей сигналов этилена и ауксина в регуляции проникновения корней у проростков томатов.
Рисунок 5.Корреляция между ингибированием рецепторов этилена, проникновением в корни и уровнем реакции на ауксин в кончиках корней томатов. A. Возобновление проникновения корней после удаления 1-MCP. Семена выращивали на Soilrite в присутствии 1-MCP в течение 3 дней. После этого 1-MCP был удален, и количество корней, проникающих в Soilrite, было подсчитано в моменты времени, указанные на оси абсцисс.B. Динамика экспрессии DR5 :: GUS в кончиках корней после удаления 1-MCP. Сеянцы выращивали без 1-MCP (верхний ряд) или в присутствии 1-MCP (нижний ряд) сразу после посева. Через 3 дня 1-MCP был удален, и после этого кончики корней были исследованы на предмет DR5 :: GUS в указанные интервалы времени. C. Одновременное применение различных ауксинов, 1-NAA, 2,4-D и IAA вместе с 1-MCP блокирует ингибирующий эффект 1-MCP на проникновение в корень в зависимости от дозы.Проникновение корней контролировали через 5 дней после посева. Звездочки указывают на статистически значимые различия между обработанными и необработанными корнями в каждой группе ( P <0,005). [См. Онлайн-статью для цветной версии этого рисунка.]
Для дальнейшего изучения этой модели мы исследовали, сможет ли экзогенно применяемый ауксин противодействовать потере проникновения в корень в присутствии 1-MCP. Проращивание семян в присутствии 1-MCP вместе с различными ауксинами, нафтилуксусной кислотой (NAA), 2,4-дихлорфеноксиуксусной кислотой (2,4-D) и IAA показало, что добавление ауксина значительно стимулировало проникновение корней. в Soilrite в зависимости от концентрации (рис.5С). Среди трех протестированных ауксинов NAA оказалась наиболее эффективной в восстановлении проникновения корней, что привело к проникновению более 50% при концентрации 0,5 мкм. Применение 2,4-Д восстановило проникновение корней при пороговой концентрации с максимальным эффектом в диапазоне от 2,5 до 5 мкм. Напротив, ИУК показала постепенное восстановление проникновения корней при увеличении концентрации. Поскольку применение 1-MCP стимулирует удлинение корня, а нанесение ауксина ингибирует удлинение корня, мы исследовали, противодействует ли ауксин эффекту 1-MCP на рост корня.Для всех ауксинов, NAA, IAA (данные не показаны) и 2,4-D (дополнительный рисунок S2, A и B), применение ауксина уменьшало удлинение корня у проростков, обработанных 1-MCP, и в то же время способствовал проникновению в корни дозозависимым образом (рис. 5C). Применение ауксина также восстановило проникновение в корни у мутанта Nr (дополнительный рис. S2C), аналогично обращению ауксинами эффекта 1-MCP. Различный эффект различных ауксинов на восстановление проникновения корней может быть связан с их проницаемостью для клеток.Хотя NAA проникает в клетки посредством диффузии через плазматическую мембрану, считается, что и IAA, и 2,4-D требуют активного транспорта для проникновения в клетку (Kramer and Bennett, 2006). Наблюдение за тем, что экзогенно поставляемый ауксин способен противодействовать индуцированному 1-МСР ингибированию проникновения в корень, предполагает, что 1-МСР действует за счет снижения уровня ауксина в кончике корня.
Транспортный мутант ауксина демонстрирует нормальное проникновение в корень в присутствии 1-MCP
Мы предположили, что если ингибирование проникновения в корень 1-MCP было связано с уменьшением поступления ауксина, то усиленный полярный транспортный мутант ауксина у томатов должен быть в состоянии противостоять индуцированному 1-MCP ингибированию проникновения в корень за счет более высоких уровней ауксина в кончике корня.Мутант томата pct1-2 обнаруживает почти в 3 раза более быстрый полярный транспорт ауксина, который связан с повышенной экспрессией переносчика ауксина PIN1 в корнях (Al-Hammadi et al., 2003; Kharshiing et al., 2010). В соответствии с нашей гипотезой, покадровая визуализация проникновения корней в pct1-2 продемонстрировала, что, в отличие от дикого типа, корни pct1-2 имели почти нормальное проникновение в Soilrite в присутствии 1-MCP, с только редкие всходы с воздушным корнем (рис.6А; Дополнительный фильм S3). Чтобы дополнительно подтвердить, что мутация pct1-2 противодействует действию 1-MCP из-за увеличения полярного транспорта ауксина, мы проверили, действует ли ингибитор транспорта ауксина 2,3,5-трийодбензойная кислота (TIBA; Dhonukshe et al., 2008). Способен снизить сопротивление 1-MCP pct1-2 при проникновении в корень. Мы наблюдали, что проростки pct1-2 были менее устойчивы к 1-MCP, когда его применяли одновременно с TIBA, показывая почти 50% ингибирование проникновения корней (рис.6, А и Б; Дополнительный фильм S4). Ингибитор притока ауксина 3-хлор-4-гидроксифенилуксусная кислота (CHPAA; Parry et al., 2001) также была эффективна в снижении устойчивости pct1-2 к 1-MCP в отношении проникновения в корень (рис. 6C). Обращение резистентности к 1-МСР в pct1-2 путем применения ингибиторов транспорта ауксина показывает, что устойчивость этого мутанта к 1-МСР является результатом, по крайней мере частично, усиленного полярного транспорта ауксина.
Рисунок 6.Мутант pct1-2 показывает нормальное проникновение в корни в присутствии 1-MCP.А. Фенотипы проростков pct1-2 , выращенных в присутствии или в отсутствие 1-MCP в течение 7 дней. Обратите внимание, что у сеянцев pct1-2 видны только случайные воздушные корни (стрелки) в присутствии 1-MCP. Обработанные TIBA (1 мкм) pct1-2 корни обычно входят в Soilrite. Однако, когда TIBA применяется одновременно с 1-MCP, почти 50% корней pct1-2 не проникают и демонстрируют воздушные петли (стрелки). B. Количественная оценка проникновения корней в проростках pct1-2 без или после обработки 1-MCP (M), TIBA (T) и 1-MCP плюс TIBA (T + M).C. Количественная оценка эффекта ингибитора притока ауксина CHPAA на проникновение в корни у проростков pct1-2 . Обратите внимание, что одновременное применение CHPAA вместе с 1-MCP ингибирует проникновение в корни у проростков pct1-2 дозозависимым образом. D, Сравнение транспорта ауксина в корнях дикого типа (WT) и pct1-2 корней. Акропетальный и базипетальный транспорт [ 3 H] IAA измеряли через 2 дня после прорастания у контрольных и обработанных 1-MCP проростков, как описано в разделе «Материалы и методы», в трех независимых экспериментах с 10-15 корнями на образец.E. Сравнение экспрессии DR5 :: GUS в кончиках корней проростков дикого типа и pct1-2 , которые были выращены на Soilrite в отсутствие или в присутствии обработки, как указано. Условия роста в пунктах A – C и E были такими же, как на фиг. 2. Звездочки и решетки указывают на статистически значимые различия в ответе на лечение, как указано, или различия между диким типом и мутантом pct1-2 , соответственно ( P <0,005; n = 15–20 на группу).[Цветную версию этого рисунка см. В статье в Интернете.]
Для дальнейшего изучения гипотезы о том, что 1-MCP ингибирует проникновение в корни за счет снижения транспорта ауксина, мы провели прямые измерения транспорта ауксина в корнях дикого типа и pct1-2 корней после обработка рассады 1-МКП. В соответствии с нашими более ранними результатами, в которых мы наблюдали усиление полярного транспорта ауксина в pct1-2 гипокотилей (Al-Hammadi et al., 2003), мы обнаружили более высокий базипетальный, а также акропетальный транспорт ауксина в корнях pct1-2 по сравнению с корнями. дикий тип.Как у дикого типа, так и у pct1-2 , применение 1-MCP вызывало снижение полярного транспорта ауксина; однако обработанные 1-МСР корни pct1-2 все еще демонстрировали повышенный транспорт ауксина по сравнению с необработанными корнями дикого типа (фиг. 6D). Эти наблюдения предполагают, что усиленный полярный транспорт ауксина в мутанте pct1-2 увеличивает уровень ауксина в кончике корня, что противодействует действию 1-MCP.
Дальнейший анализ показал, что активность DR5 была увеличена в pct1-2 кончиках корней по сравнению с диким типом (рис.6E). Хотя воздействие 1-MCP снижало активность DR5 как в кончиках корня дикого типа, так и в кончиках корня pct1-2 , экспрессия DR5 все еще оставалась намного выше в кончиках корня pct1-2 по сравнению с диким типом. Применение ТИБА вместе с 1-MCP вызывало резкое снижение активности DR5 как у корней дикого типа, так и у кончиков корней pct1-2 (фиг. 6E). Это было связано с ингибированием проникновения корней в pct1-2 (фиг. 6, A и B). Взятые вместе, приведенные выше результаты согласуются с гипотезой о том, что усиленный транспорт ауксина в мутанте pct1-2 противодействует способности 1-MCP ингибировать проникновение в корень, и они поддерживают взаимодействие между ауксином и этиленом в регулировании этого физиологического состояния. отклик.
ОБСУЖДЕНИЕ
Ингибирование действия этилена препятствует проникновению корней и вызывает воздушный рост корней
Для выполнения важной задачи обеспечения закрепления и обеспечения питательными веществами растений корни должны непрерывно расти и проникать в почву. Одна из основных сельскохозяйственных проблем — механическое сопротивление росту корней в плотных почвах. Хотя ученые-растениеводы исследовали феномен проникновения корней через плотную почву в различных условиях окружающей среды, имеется очень ограниченная информация о молекулярных процессах, лежащих в основе этого явления (Clark et al., 2003). Исследования показали, что определенные морфологические изменения, такие как увеличение диаметра корня за верхушкой корня при столкновении с более высоким механическим сопротивлением, могут быть связаны со способностью корней проникать в твердые почвы (Wilson et al., 1977). Физиологические данные показали, что при столкновении с механическим сопротивлением локализованное увеличение синтеза этилена вызывает увеличение диаметра корня (Le et al., 2001). В соответствии с этой точкой зрения, корни мутанта кукурузы с дефектом активности АСС-синтазы, ключевого фермента, регулирующего синтез этилена, показали снижение роста в почве, что свидетельствует о важности этилена в преодолении физической устойчивости (Gallie et al., 2009). Целью нашего исследования было проанализировать роль этилена в регуляции проникновения корней и возможную роль ауксина в содействии этой реакции у проростков томатов.
Неспособность первичного корня томата проникать в среду в условиях ингибирования рецепторов этилена либо за счет использования специфических ингибиторов (Zacarias and Reid, 1992), либо из-за мутаций, таких как в гене Nr (Clark et al., 1999), демонстрирует важную роль этилена в регулировании проникновения корней.В соответствии с предыдущими исследованиями, корни проростков томатов, выращенных в присутствии 1-МСР, ингибитора рецепторов этилена (Sisler et al., 1996; Binder and Bleecker, 2003; Feng et al., 2004; Sisler, 2006), не удалось проникнуть в Soilrite. Точно так же корни устойчивого к этилену мутанта Nr также не смогли проникнуть в Soilrite, ответ, который усугублялся применением 1-MCP. Эффект 1-MCP не ограничивался ингибированием проникновения корней; он также повлиял на рост как гипокотилей, так и корней со стимуляцией роста корней и ингибированием удлинения гипокотилей.Поскольку этилен способствует удлинению гипокотилей у выращенных на свету проростков (Smalle et al., 1997; Vandenbussche et al., 2003a, 2003b) и ингибирует удлинение корня (Smalle and van der Straeten, 1997; Buer et al., 2006), наблюдаемые ингибирование длины гипокотиля и увеличение длины корня согласуется с ингибированием действия этилена у проростков, обработанных 1-MCP. Увеличение количества боковых корней у проростков, обработанных 1-MCP, также напоминало увеличение числа проростков Nr (Negi et al., 2010), дополнительно подтверждая, что 1-MCP действует, влияя на реакцию этилена. Кроме того, замечательный фенотип воздушного роста корней, который был связан с отсутствием проникновения корней в присутствии 1-MCP, наблюдался у однодольных растений (рис и пшеница) и двудольных растений (табак и салат), что указывает на существенную роль этилена, способствующего проникновению корней в питательную среду, эволюционно сохраняется.
Эксперименты с переносом проростков томатов в атмосферу с 1-MCP через различные интервалы времени после посева показали, что сигнальный путь, обеспечивающий проникновение корней, инициируется на очень ранней стадии прорастания семян, возможно, до появления корешка из семян. Пальто.Как только корешок выходит из семенной оболочки и корень проникает в Soilrite, влияние 1-MCP на проникновение корня уменьшается. Однако способность 1-MCP ингибировать проникновение в корень не связана с ингибированием прорастания корешка из семян или с уменьшением выделения этилена на ранней стадии прорастания корешка.
Поскольку кончик корня играет важную роль в определении направления силы тяжести, а также механического взаимодействия между корнем и почвой (Massa and Gilroy, 2003; Bengough et al., 2006), а этилен модулирует гравитропизм корней у Arabidopsis (Buer et al., 2006), можно предположить, что 1-MCP влияет на проникновение в корни, ингибируя гравиответ или механочувствительность корней. Однако воздушный рост корней, по-видимому, не является результатом потери гравитропной реакции, поскольку кончики корней проростков, обработанных 1-MCP, оставались положительно гравитропными. Покадровая съемка постоянно показывала, что у этих проростков кончики корней оставались в тесном контакте с поверхностью Soilrite, а отсутствие проникновения заставляло корни расти в воздухе, образуя петли.Сходным образом идея о том, что 1-MCP может ингибировать проникновение в корни за счет снижения механочувствительности, не подтверждается наблюдением, что потенциальный связанный с механовосприниманием транскрипт, SlMCA1 , активируется в корнях, обработанных 1-MCP. Тот факт, что ингибирующее действие 1-MCP на проникновение корней было обращено этиленом, показывает, что 1-MCP не навсегда снижает способность кончика корня проникать в почву. Возобновление проникновения корней в Soilrite при изъятии 1-MCP наблюдалось через 12 часов, а через 48 часов все корни проникли в Soilrite.Поскольку считается, что 1-MCP образует стабильную связь с рецепторами этилена (Sisler et al., 1996; Binder and Bleecker, 2003; Feng et al., 2004), период времени от 12 до 48 часов может отражать время требуется для восстановления рецептора и / или синтеза новых рецепторов этилена, чтобы повторно инициировать реакцию этилена и процесс проникновения в корень. По сути, эти наблюдения указывают на ключевую роль этилена в регулировании проникновения корней.
Проникновение в корни, вероятно, регулируется перекрестным разговором по этилен-ауксину
У Arabidopsis ауксин и этилен обладают синергетическим действием на несколько реакций роста и развития корней, включая инициирование и рост корневых волосков (Pitts et al., 1998), гравитропизм (Buer et al., 2006) и удлинение корня (Pickett et al., 1990; Rahman et al., 2001; Swarup et al., 2002; Rzicka et al., 2007). Поэтому неудивительно, что ингибирование проникновения 1-МСР в корни сопровождалось изменениями экспрессии генов ответа на этилен и ауксин. Это говорит о том, что два гормона тесно взаимодействуют, регулируя проникновение корней. Интересно, что 1-МСР по-разному влиял на экспрессию генов ETR в корнях и гипокотилях.В то время как экспрессия транскриптов рецепторов ETR2 , -4 , -5 и — 6 увеличивалась в корнях, экспрессия ETR3 , -4 , -5 и — 6 уменьшилось в гипокотилях. Ранее сообщалось, что 1-MCP снижает экспрессию ETR4 / 5/6 , но оказывает маргинальное влияние на экспрессию ETR1 / 2 и Nr ( ETR3 ) в плодах томата на стадии разрушения, в корреляция с задержкой созревания плодов (Tassoni et al., 2006). Следовательно, дифференциальный ответ рецепторов ETR на обработку 1-MCP может быть связан с их дифференциальным участием в различных ответах развития. Однако повышение уровней экспрессии некоторых рецепторов ETR в корнях, обработанных 1-MCP, также может быть связано с компенсаторным ответом, возникающим в результате ингибирования рецептора на уровне белка, чтобы помочь поддерживать гомеостаз и действие этилена. Компенсаторное взаимодействие между различными рецепторами наблюдалось у томатов, где снижение экспрессии рецепторов ETR1 / 3 было компенсировано увеличением экспрессии ETR4 (Tieman et al., 2000).
У Arabidopsis генетические данные свидетельствуют о том, что этилен подавляет рост корней частично за счет активации биосинтеза ауксина (Stepanova et al., 2005). Следовательно, возможно, что ингибирование передачи сигналов этилена 1-MCP, в свою очередь, приводит к снижению уровня ауксина в корнях. В соответствии с этой точкой зрения, наши наблюдения продемонстрировали, что обработка 1-MCP была отчетливо связана со снижением активности репортера ауксинового ответа DR5 в кончиках корней, предполагая, что неспособность корней проникать в почву была связана с снижение ауксинового ответа в верхушке корня.Более того, мы наблюдали, что активность DR5 обратимо восстанавливалась после отмены 1-MCP, параллельно с возобновлением проникновения в корень. Это убедительно подтверждает причинную связь между передачей сигналов ауксина и проникновением в корни, которое модулируется этиленом. Связь между передачей сигналов ауксина и проникновением в корни дополнительно подтверждается наблюдением, что одновременное применение различных ауксинов вместе с 1-MCP к прорастающим семенам постепенно обращало ингибирующее действие 1-MCP на проникновение в корни в зависимости от дозы.Более высокая относительная эффективность NAA в противодействии действию 1-MCP по сравнению с IAA и 2,4-D согласуется с более высокой клеточной проницаемостью этой формы ауксина (Marchant et al., 1999). Таким образом, эти результаты подтверждают причинную связь между передачей сигналов ауксина и проникновением в корни, которое модулируется этиленом.
В соответствии с перекрестным взаимодействием этилена и ауксина, участвующим в способности корней проникать в почву, мы также наблюдали, что уровни транскриптов SlIAA3 и SlIAA9 модулировались во время индуцированного 1-MCP ингибирования проникновения корней.Снижение экспрессии ауксин-чувствительного гена в корнях, обработанных 1-MCP, которое отслеживается по снижению активности DR5, может приводить к повышенной экспрессии компонентов, ингибирующих действие ауксина. Передача сигналов ауксина у высших растений модулируется белками Aux / IAA, которые действуют как репрессоры транскрипции регулируемых ауксином генов, включая их собственные, и ауксин регулирует их экспрессию как на уровне транскрипции, так и на уровне стабильности белка (Woodward and Bartel, 2005). Функции SlIAA3 и SlIAA9 были продемонстрированы в развитии плодов и вегетативных побегов (Wang et al., 2005; Chaabouni et al., 2009). Напр., Подавление транскрипта SlIAA3 в томате с использованием РНК-интерференции приводит к появлению апикального крючка с преувеличенной кривизной у выращенных в темноте растений и уменьшению эпинастии черешка у выращенных на свету растений (Chaabouni et al., 2009). Подавление транскрипта SlIAA9 приводит к гиперчувствительности к ауксину, и у трансгенных растений появляются простые листья и развиваются партенокарпические плоды (Wang et al., 2005). Поскольку транскрипты SlIAA3 и SlIAA9 активируются при ингибировании проникновения в корень с помощью 1-MCP, эти компоненты передачи сигналов ауксина могут действовать как негативные регуляторы проникновения в корень.Взятые вместе, наши результаты подтверждают вероятность перекрестной передачи сигналов этилена и ауксина в регуляции проникновения корней у проростков томатов.
Этилен Повышение полярного транспорта ауксина участвует в проникновении в корень
Как этилен может влиять на уровень ауксина и / или реакцию во время проникновения в корень? Предыдущие сообщения показали, что этилен может стимулировать как транспорт ауксина, так и биосинтез в корнях (Stepanova et al., 2005, 2007; Růzicka et al., 2007; Swarup et al., 2007). Корни мутанта Nr томата, который демонстрирует пониженное восприятие этилена, как у проростков, обработанных 1-MCP, демонстрируют сниженный полярный транспорт ауксина, тогда как мутант Epi с избыточным продуцированием этилена демонстрирует повышенный транспорт ауксина (Negi et al., 2010). У Arabidopsis этилен регулирует транскрипцию нескольких переносчиков ауксина, включая PIN1, PIN2 и AUX1 (Růzicka et al., 2007), тем самым усиливая транспорт ауксина в корнях. Следовательно, наблюдаемое снижение активности DR5 в корнях, обработанных 1-MCP, может быть результатом снижения биосинтеза ауксина, полярного транспорта ауксина или комбинации обоих этих процессов.Наши данные о том, что применение 1-MCP снижает транспорт акропетального и базипетального ауксина в корнях томатов дикого типа, подтверждают, что этилен действует на проникновение в корни, по крайней мере частично, за счет увеличения полярного транспорта ауксина. Эта точка зрения была дополнительно подтверждена наблюдением, что мутант pct1-2 , который имеет почти в 3 раза более высокий транспорт ауксина, чем дикий тип (Al-Hammadi et al., 2003), показал почти нормальное проникновение корней в Soilrite в наличие 1-МКП. Более высокий уровень ауксина в корнях pct1-2 может противодействовать ингибирующему эффекту 1-MCP на проникновение в корни.Хотя обработка 1-MCP уменьшала транспорт ауксина в корнях pct1-2 , полярный транспорт ауксина все еще был значительно выше в обработанных 1-MCP корнях pct1-2 , чем в необработанных корнях дикого типа. Тот факт, что активность DR5 в корнях pct1-2 была значительно выше, чем в корнях дикого типа, даже в присутствии 1-MCP, поддерживает мнение о том, что повышенный уровень ауксина способствовал проникновению в корни pct1-2 саженцы. По сути, эти результаты предполагают, что повышение уровня ауксина в корнях pct1-2 преодолевает ингибирующее действие 1-MCP на проникновение в корни.
Если способность корней pct1-2 нормально проникать через Soilrite в присутствии 1-MCP обусловлена повышенным транспортом ауксина, то при ингибировании полярного транспорта ауксина мутант pct1-2 должен показать ингибированный корень. проникновение аналогично тому, которое наблюдается у дикого типа. Сообщалось, что несколько ингибиторов ингибируют полярный транспорт ауксина, такие как TIBA, который препятствует перемещению везикул переносчиков ауксина (Dhonukshe et al., 2008), и CHPAA, ингибитор притока ауксина (Parry et al., 2001). В соответствии с тем, что pct1-2 способен противостоять ингибирующему эффекту 1-MCP на проникновение в корень из-за более высокого транспорта ауксина, одновременное применение TIBA или CHPAA вместе с 1-MCP частично ингибировало проникновение в корень в pct1-2 , что приводит к образованию четких воздушных петель. Одновременно активность DR5 также снизилась в pct1-2 кончиках корней, потенциально обнаруживая общее снижение экспрессии ауксин-чувствительного гена.
Таким образом, наши результаты показывают, что совместное действие этилена и ауксина, вероятно, регулирует проникновение корней томатов в почву.Хотя молекулярные механизмы, регулирующие проникновение корней, еще предстоит расшифровать, у томатов, по-видимому, существует обязательное требование действия этилена для осуществления проникновения корней. После этого передача сигналов этилена модулирует синтез и / или транспорт ауксина, чтобы запустить путь, ответственный за проникновение корней в почву. Хотя наше исследование показывает, что транспорт ауксина играет ключевую роль в регулировании проникновения в корни, возможно, что перекрестные помехи между этиленом и ауксином могут происходить на многих разных уровнях, включая модуляцию чувствительности к ауксину и его накопления.В будущем более подробный анализ взаимодействия между этими двумя гормонами, а также другими регуляторными молекулами может раскрыть точный механизм, управляющий процессом проникновения в корни.
МАТЕРИАЛЫ И МЕТОДЫ
Растительные материалы и условия роста
Томаты дикого типа ( Solanum lycopersicum ), мутант Nr и мутант pct1-2 (Madishetty et al., 2006) семена, все в В этом исследовании использовались данные Эйлсы Крейг.Мутант pct1-2 был скрещен с трансгенной линией DR5 :: GUS (Dubrovsky et al., 2008), и были использованы гомозиготные проростки pct1-2 с GUS-активностью в поколении F4. Поверхность семян стерилизовали 20% (об. / Об.) Гипохлоритом натрия в течение 10 минут, промывали и непосредственно высевали (если не указано иное) на поверхность влажного Soilrite (смесь вспученного перлита для садоводства, ирландского торфяного мха и расслоенного мха). вермикулит в равных пропорциях; Karnataka Explosive) в прозрачных пластиковых ящиках для проращивания (9.5 см в длину × 9,5 см в ширину × 5 см в высоту). Растения выращивали при постоянном белом свете (100 мкмоль м -2 с -1 ) в течение 7 дней, если не указано иное. Имеющиеся в продаже семена риса ( Oryza sativa ), пшеницы ( Triticum aestivum ), табака ( Nicotiana tabacum ) и салата ( Lactuca sativa ) стерилизовали и выращивали на Soilrite, следуя той же процедуре. Для опытов с вертикальным ростом проростков после стерилизации семена проращивали на 1.2% (мас. / Об.) Агар на чашках Петри. Если не указано иное, все эксперименты повторяли не менее трех раз.
Обработка 1-MCP
Если не указано иное, 30 мг порошка, высвобождающего 1-MCP (0,14% активного ингредиента по весу; SmartFresh [Rohm and Haas]), растворяли в 5 мл воды в пластиковом флаконе для получения конечная концентрация газа 2 мкл л –1 1-MCP (Tassoni et al., 2006). Пробирку приклеивали клейкой лентой к стенке ящика для проращивания, и ящики немедленно плотно закрывали герметизирующей лентой.В комбинированных обработках вместе с 1-MCP добавляли ингибитор транспорта ауксина, 1 мкМ TIBA (Sigma-Aldrich) или CHPAA (Sigma-Aldrich), и семена высевали на Soilrite, содержащем соответствующие растворы. Флакон, содержащий 1-MCP, был прикреплен лентой к бокам ящиков для проращивания, и ящики были плотно закрыты. Для одновременного воздействия этилена и 1-MCP флакон с 1-MCP помещали в ящики для проращивания, как описано выше. Кроме того, через резиновую перегородку в герметично закрытый бокс вводили различные количества газообразного этилена.Частоту проникновения корней определяли через 7 дней после посева.
Чтобы изучить влияние 1-MCP на формирование боковых корней, проростки выращивали на чашках Петри (глубиной 14 см) на 1,2% (мас. / Об.) Агаре в присутствии или в отсутствие 1-MCP. После посева чашки Петри ориентировали вертикально на 9 дней. Флакон, содержащий 1-MCP, был прикреплен лентой к вертикальной крышке чашек Петри. Для исследования влияния 1-MCP на прорастание семена высевали на агар (0,8%, мас. / Об.) В присутствии или в отсутствие 1-MCP.Начало прорастания оценивали путем визуального наблюдения за появлением корешка из семенной оболочки через 12-часовые интервалы после посева. Чтобы определить момент времени начала действия 1-MCP на проникновение корней, семена высевали на Soilrite. Флаконы, содержащие 1-MCP, помещали в ящики с 24-часовыми интервалами с 0 дня до 4 дней после посева. Контрольные проростки выращивали в течение 5 дней без обработки 1-MCP. Количество корней, проникающих в Soilrite, подсчитывали через 5 дней после посева.Для определения времени возобновления проникновения корней после удаления 1-MCP проростки выращивали в присутствии 1-MCP в течение 3 дней после посева. После этого флакон с 1-MCP был удален, и количество первичных корней, попавших в Soilrite, было подсчитано с 12-часовыми интервалами.
Определение эволюции этилена
Для измерения выделения этилена из проростков растения выращивали в плотно закрытых коробках для проращивания. Через 24 ч после посева из флаконов через резиновую перегородку с помощью шприца удаляли 1 мл свободного пространства.Затем газовый поток вводили в газовый хроматограф, оборудованный колонкой Poropak T и пламенно-ионизационным детектором (GC-17A; Shimadzu) для определения уровней этилена. Общее выделение этилена нормировали на количество проростков и время инкубации. В каждый момент времени содержание этилена в газе свободного пространства измеряли три раза.
Окрашивание GUS
Кончики корней инкубировали в 1 мл раствора для окрашивания GUS, содержащего 10 мМ натрий-фосфатный буфер (pH 7,2), 10 мМ ЭДТА, 1 мМ 5-бром-4-хлор-3-индолил-β-d -GlcA циклогексиламмониевая соль и 1% (об. / Об.) Triton X-100.Для облегчения инфильтрации наконечники подвергали вакууму в течение 5 минут и оставляли под вакуумом на 30 минут. После этого кончики инкубировали при 37 ° C в темноте в течение 16 часов. Кончики корней очищали в течение ночи в растворе формальдегид: абсолютный спирт: уксусная кислота (2: 10: 1) и наблюдали с помощью составного микроскопа Olympus BX41, закрепленного с помощью цифровой камеры (Olympus C-7070).
Анализ экспрессии гена с помощью qRT-PCR
Кончики корней (1 см) и гипокотили (1,5 см) ниже семядолей вырезали из 5-дневных проростков, выращенных на 0.8% (мас. / Об.) Агар в ящиках для проращивания в присутствии или в отсутствие 1-MCP для облегчения сбора кончиков корней. Ткани замораживали в жидком N 2 и гомогенизировали до тонкого порошка, используя ступку и пестик. Около 100 мг гомогенизированного порошка использовали для экстракции общей РНК с помощью набора RNeasy (Qiagen) в соответствии с инструкциями производителя. Концентрацию РНК определяли путем измерения УФ-поглощения при A 260 и A 280 на спектрофотометре Nanodrop.Целостность РНК проверяли электрофорезом в агарозном геле. RT выполняли с 2 мкг общей РНК в общем объеме 20 мкл с использованием набора для синтеза кДНК (Bioline). qRT-PCR выполняли с использованием кДНК, соответствующих 5 нг общей РНК в реакционных объемах 20 мкл, с использованием SYBR Green PCR Master Mix (Takara) в системе 7300 Fast Real Time PCR (Applied Biosystems). Праймеры для гена MCA1 были сконструированы из EST томата (SGN-E 1245095), выровненного с SGN Unigene (SGN-U565830), который был получен путем BLAST-анализа последовательности гена MCA1 (At4g35920) Arabidopsis (At4g35920) против базы данных SGN Unigene ( http: // solgenomics.сеть). Ген-специфические праймеры, используемые для qRT-PCR, были разработаны с использованием программного обеспечения PRIMER3. Анализ qRT-PCR проводили, как описано ранее (Pirrello et al., 2006). Оптимальная концентрация праймера составляла 280 нм для ген-специфических праймеров. Чтобы определить относительные кратные различия для каждого образца в каждом эксперименте, значения порога цикла (Ct) для генов SlERF2 и SlMAN2 были нормализованы к значению Ct для SlAct51 и рассчитаны относительно калибратора с использованием уравнения 2 −ΔΔCt (Ливак, Шмитген, 2001).
Последовательности праймеров были следующими: для 18SrRNA, прямая (FP), 5′-GCGACGCATCATTCAAATTTC-3 ‘и обратная (RP), 5′-TCCGGAATCGAACCCTAATTC-3’; для ETR1 , FP, 5′-GTTGCCTGCTGACGACTTGC-3 ‘и RP, 5′-GCACCGAACTGCACAAGAACC-3’; для ETR2 , FP, 5′-TGCTGCCATCTTGGAGGAATC-3 ‘и RP, 5′-GAAAGTGCAACCACCGCATG-3’; для ETR3 , FP, 5′-CGCAGATCAGGTTGCTGTCG-3 ‘и RP, 5′-TGGGCGTTCTCATTTCATGG-3’; для ETR4 , FP, 5′-AGCCAGAGGGGACCATGTTG-3 ‘и RP, 5′-CCCAAGAACGACAGCCATGC-3’; для ETR5 , FP, 5′-ATTCGGATGCCACTGCTTCG-3 ‘и RP, 5′-GGATAAAGCCACAGCCACCTG-3’; для ETR6 , FP, 5′-CCCCTGCTCCTCCAACATACG-3 ‘и RP, 5′-ATCCGACAATCACAGCCATGC-3’; для IAA9 , FP, 5′-GATTGTTCGTCGGTGGACAGC-3 ‘и RP, 5′-AAGGGTGCAAGGGGAAGAGC-3’; для IAA3 , FP, 5′-GCATGGATGGAGCACCTTATTTG-3 ‘и RP, 5′-TGGTACATCACCAACAAGCATCC-3’; и для MCA1, подобного , FP, 5′-GCCCTCCTGCTGATCTGTGC-3 ‘и RP, 5′-CCCAATTAGCCTCACCGCATC-3’.
Анализ транспорта ауксина
Полярный транспорт ауксина измеряли с использованием метода Negi et al. (2010) и Lewis and Muday (2009) с небольшими изменениями. Для анализа акропетального транспорта семена проращивали на промокательной бумаге в присутствии или в отсутствие 1-MCP, и после появления корешка их переносили на чашки с агаром. Через 48 часов агарную линию 100 нм [ 3 H] IAA наслаивали непосредственно под выровненными стыками корень-побег, и проростки инкубировали на свету в перевернутом положении в течение 18 часов.В конце лечения иссекали апикальные 5 мм каждого кончика корня и использовали их для измерения радиоактивности. Десять кончиков корней на обработку объединяли в 2,5 мл сцинтилляционной жидкости и измеряли радиоактивность в течение 2 минут в жидкостном сцинтилляционном счетчике. Для измерения базипетального транспорта ауксина проростки, выращенные, как описано выше, обрабатывали путем нанесения агаровой линии 100 нм [ 3 H] IAA, прилегающей к верхушке корня, и проростки инкубировали на свету в течение 5 часов. После обработки апикальные 2 мм корня вырезали и отбрасывали, а 5-миллиметровые сегменты над верхушкой отбирали для количественного определения радиоактивной ИУК.
Анализ корневого гравитропизма
Анализ корневого гравитропизма контрольных и обработанных 1-MCP проростков выполняли, как описано Al-Hammadi et al. (2003).
Дополнительные данные
Следующие материалы доступны в онлайн-версии этой статьи.
Благодарности
Мы благодарим доктора Сангиту Неги за полезные обсуждения и Рома и Хааса за любезный подарок SmartFresh.
Сноски
Автор отвечает за распространение материалов, составляющих выводы, представленные в этой статье, в соответствии с политикой, описанной в Инструкциях для авторов (www.plantphysiol.org): Рамешвар Шарма (rameshwar.sharma {at} gmail.com).
www.plantphysiol.org/cgi/doi/10.1104/pp.111.177014
↵1 Эта работа была поддержана Международным агентством по атомной энергии, Департаментом биотехнологии, Нью-Дели, Индия (гранты для RS и YS), Комиссией по университетским грантам, Нью-Дели, Департаментом биотехнологии — Центром образования и исследований в области биологии и биотехнологии (стипендия PS), стипендией Университета Хайдарабада для N.S., и Программой конкурсных грантов Национальной исследовательской инициативы Министерства сельского хозяйства США (грант № 2006–03434, выданный M.G.I.).
↵ [C] Некоторые рисунки в этой статье отображаются в Интернете в цвете, а в печатном издании — в черно-белом.
↵ [OA] Статьи в открытом доступе можно просматривать в Интернете без подписки.
↵ [W] Онлайн-версия этой статьи содержит данные только для Интернета.
- Поступила 25.03.2011.
- Принято 10 мая 2011 г.
- Опубликовано 12 мая 2011 г.
Оптимизация корневых систем сельскохозяйственных культур — Новый Южный Уэльс | Информационные бюллетени
Ключевые моменты
- Увеличение количества и глубины корней сельскохозяйственных культур означает, что растения могут потреблять больше питательных веществ и воды.
- Поддерживайте pH выше 5,2 в верхнем слое почвы и 4,8 в подпочве, чтобы питательные вещества оставались доступными.
- Другими ограничениями недр, на которые следует обратить внимание, являются уплотнение, соленость и соленость.
- Улучшение структуры почвы может снизить риск и серьезность корневых заболеваний.
Увеличение количества и глубины корней
Почвенные условия влияют на развитие корневой системы сельскохозяйственных культур. Один из способов оптимизировать усвоение растениями запасов питательных веществ в почве — это максимизировать площадь, которую корни сельскохозяйственных культур могут контактировать с почвой, и обеспечить, чтобы они могли следить за питательными веществами и поливать профиль. Это особенно важно для выщелачиваемых питательных веществ, таких как азот и сера, и питательных веществ со значительными запасами в недрах. Обилие и глубину корней сельскохозяйственных культур можно увеличить, распознавая и улучшая условия, которые препятствуют развитию или продуктивности корней.
pH почвы
Доступность питательных веществ оптимизирована в почвах с уровнем pH верхнего слоя почвы в диапазоне pH 5,2–7,5 (измеряется в CaCl 2 ). Поддержание pH выше 4,8 в подпочвенном слое будет способствовать росту корней в подпочвенных горизонтах. Когда pH падает ниже 4,8, корневые системы, как правило, не проникают в кислую почву, и поэтому они не могут получить доступ к воде и питательным веществам из более глубоких слоев почвы. Сельскохозяйственная известь эффективна для снижения кислотности почвы и поддержания соответствующего pH (см. Информационные бюллетени «Кислотность почвы» и «Анализ химических показателей»).Построение органического вещества почвы улучшает буферный уровень pH и, таким образом, помогает почвенным организмам и культурам справляться с кислыми условиями.
Уплотнение грунта
Уплотнение почвы может уменьшить распространение корневой системы сельскохозяйственных культур за счет замедления и ограничения проникновения корней (рисунок 1). Плохая аэрация приводит к анаэробным условиям, при которых рост корней и биологическая активность ограничиваются чаще и на более длительные периоды по сравнению с аналогичной, но лучше аэрированной почвой. Это может означать, что корни не могут получить доступ к питательным веществам, таким как азот, из недр, и поэтому эффективность использования азота низкая.
Рис. 1: Отчетливый уплотненный слой в супеси; обратите внимание на трещины в твердой посуде, через которые преимущественно прорастают корни.
Глубокий рыхление может быть эффективным методом уменьшения уплотнения грунта, когда осмотр профиля грунта показывает уплотненные слои (особенно слои, которые являются непрерывными и будут препятствовать проникновению корней, см. Информационный бюллетень «Уплотнение грунта»). Эффект разрыва является временным, и, если не будут внесены изменения в руководство, направленные на уменьшение продолжающегося уплотнения, уплотнение произойдет снова.Некоторые усовершенствованные методы — это контролируемое движение или выращивание глубоко укоренившихся многолетних растений.
Sodicity
Содичность — это когда доля натрия по сравнению с другими обменными катионами в почве слишком высока. В поверхностных почвах содовость вызывает диспергирование и эрозию почвы, но баланс обменных катионов можно скорректировать с помощью гипса, извести и органических веществ. С глубиной все труднее устранять нелестные отношения.
Химический состав натриевых грунтов может ограничивать рост и продуктивность корней, ограничивая потребление воды и некоторых питательных веществ, позволяя усваивать другие питательные вещества сверх потребностей растений.Многие натриевые грунты практически непроницаемы. После дождя посевы подвергаются как переувлажнению поверхностного слоя почвы, так и стрессу влаги в подпочве из-за недостаточного проникновения воды в почву.
Глубокое рыхление может способствовать проникновению и росту корней в краткосрочной перспективе, но также может привести к тому, что заполнители снова станут хуже, чем до рыхления. Применение гипса с глубоким рыхлением снижает этот риск.
Соленость
Засоленная почва содержит высокие концентрации растворимых солей, которые ограничивают рост большинства растений.Соль — это любое химическое соединение, в котором катион (положительный ион) уравновешивается анионом (отрицательный ион). Самая распространенная соль — хлорид натрия. Многие почвенные добавки, включая гипс, известь и многие удобрения, также являются солями. Засоление снижает способность растений извлекать воду из почвы и может вызывать токсичность определенных ионов.
Засоление почв происходит из-за взаимодействия с грунтовыми или поливными водами. Если грунтовые воды поднимаются до уровня в пределах 2 м от поверхности почвы, вода может подняться на поверхность за счет капиллярного действия.Когда это происходит, вода может приносить соль в корневую зону, а когда вода испаряется на поверхности почвы, остаются концентрированные соли (рис. 2).
Рис. 2: Ошпаривание из-за солености. Фото: Грэм Джонсон © правительство Нового Южного Уэльса.
Соли в почвенных добавках вряд ли будут применяться в достаточно больших количествах, чтобы создать проблему засоления, но там, где засоление уже существует, эти добавки соли могут иметь отношение к выражению проблем засоления.
Основные стратегии управления и предотвращения засоления почв направлены на поддержание глубины грунтовых вод на глубине менее 2 м за счет максимального использования воды растениями и на максимально эффективное орошение (см. Информационный бюллетень «Засоление»). Выбор устойчивых сортов сельскохозяйственных культур важен там, где уже присутствует засоление.
Болезни корней растений
Болезни, поражающие корни растений, могут существенно снизить способность растения получать воду и питательные вещества из почвы. Хорошая структура почвы и здоровая биология почвы помогают снизить риск и степень распространения корневых заболеваний сельскохозяйственных культур.Переувлажнение или водный стресс способствуют развитию многих корневых заболеваний. Хорошая структура позволяет отводить излишки воды и пропускать кислород в профиль почвы во влажных условиях, максимально увеличивает доступную воду для растений в сухих условиях и обеспечивает подходящие условия для биологии почвы. Здоровые популяции разнообразных почвенных организмов снижают способность болезнетворных организмов распространяться и доминировать в почве, и это может уменьшить повреждение сельскохозяйственных культур даже при наличии болезни.
Севообороты с видами, не являющимися хозяевами, могут сократить популяции болезнетворных организмов, если хорошо бороться с сорняками.Если сорняки не контролируются, некоторые виды могут поддерживать патоген во время перерыва в севообороте, что снижает пользу от практики. Сжигание стерни может удалить болезнетворные инокуляты над землей, но существенно не снижает присутствие болезнетворных организмов в почве. См. Информационные бюллетени «Fusarium», «Nematodes», «Rhizoctonia», «Take-all Disease», «Soil Bacteria and Fungi».
Дополнительная литература и ссылки
У Департамента первичной промышленности Нового Южного Уэльса есть информация о почвах и управлении ими (онлайн), в том числе о содности и засолении, а также информация о болезнях сельскохозяйственных культур (онлайн)
Автор: Стефани Альт (Департамент первичной промышленности Нового Южного Уэльса), 2013 г.
Национальная программа мониторинга качества почв финансируется Корпорацией исследований и развития зерновых в рамках второй инициативы по биологии почв.
Участвующие организации не несут никакой ответственности по причине небрежности или иным образом, возникшей в результате использования или разглашения этой информации или любой ее части.
Посмотреть все информационные бюллетени
Что такое сварной шов с полным проплавлением?
Как известно, дефекты техники сварки могут отрицательно сказаться на качестве свариваемого материала.При сварке материала используются различные методы. В этой статье вы узнаете о технике сварки с полным проплавлением, также известной как сварка с полным проплавлением.
Что такое сварной шов с полным проплавлением? Техника сварки с полным проплавлением включает соединение двух металлических частей без зазоров между ними. В этом типе сварного шва присадочный материал заполняет корни шва. Этот метод в основном применяется для металлических изделий с высокой нагрузочной способностью.
Метод полного проникновения дает более высокую прочность по сравнению с частичным проникновением.В результате металлические изделия с полным проплавлением идеально подходят для металлических изделий, предназначенных для работы с высокими напряжениями.
При сварке с полным или полным проплавлением получается толстый сварной шов. Сварной шов скошен и проплавлен различными сварочными проходами. Однако в соединениях с толщиной 0,20 дюйма или более проплавление достигается без скоса канавки.
Сварочный шов с полным или полным проплавлением может быть толстым сварным швом, который был скошен, а затем пропущен путем заполнения несколькими проходами, или это может быть соединение листового металла.020 ″ и проходит без фаски канавки.
Глубокое проникновение требует очень высокой плотности мощности более 1 мегаватта на квадратный метр. Полученная в результате балка обеспечивает улучшенную сварку и более прочный металлический продукт.
При сварке с полным проплавлением металлические детали плавятся встык, который полностью плавится с основным металлом. Прочность основного металла во всех соединениях одинакова. Этот шов с полным проплавлением может иметь различные конфигурации, включая конфигурации с одним и двумя клиньями.
Если вы выполнили сварку с частичным проплавлением, сплавленный металл будет иметь более низкий коэффициент сопротивления прочности по сравнению с основным металлом.
Как достигается сварка с полным проплавлением?
На качество сварки с проплавлением влияют разные факторы. Давайте посмотрим на некоторые факторы, влияющие на качество сварки.
1. Сварочный ток
Сварочный ток — главный фактор, влияющий на проплавление сварного шва. Более высокий сварочный ток приведет к более глубокому проплавлению.Противоположный случай имеет место при более низком сварочном токе.
Вы можете изменить сварочный ток, отрегулировав скорость подачи проволоки. Другими словами, желаемый ток может быть достигнут путем изменения расстояния между наконечником и металлом.
Между расстоянием между наконечником и металлом и сварочным током существует противоположная зависимость. Если увеличить расстояние, ток уменьшится. Это приведет к частичному проникновению. Напротив, если вы уменьшите расстояние от наконечника до металла, ток увеличится, что приведет к сварке с полным проплавлением.
Изменяя расстояние от наконечника до металла, вы можете контролировать ток, подаваемый при сварке металлических деталей. Обратите внимание, что в некоторых случаях вам не следует проводить сварку на определенном расстоянии в течение длительного времени. Подумайте о том, чтобы немного изменить расстояние, чтобы предотвратить прожог в случае неоднородностей в зазоре стыка.
2. Стабильность дуги
Стабильность дуги — еще один фактор, влияющий на полное проплавление шва. Для глубокого проплавления дуга должна быть стабильной. Скорость прохождения дуги напрямую влияет на степень проникновения.
Вы можете добиться большего проплавления с более высокой скоростью движения дуги. Плавление будет полным при скорости дуги 12 дюймов в минуту (ipm) или 30,5 сантиметров в минуту (cpm). При 7 изобр. / Мин или 17,8 циклов в минуту проникновение уменьшится.
Уровень проплавления также снизится, если скорость прохождения дуги превысит 17 дюймов в минуту или 43,2 дюйма в минуту. При более высоких скоростях тепло, выделяемое при сварке, не успевает расплавить область основного металла. В случае более низкой скорости дуги в области откладывается много присадочного материала.Жидкий сварочный валик спереди предотвращает более глубокое проплавление.
3. Положение резака
Положение сварочной горелки также влияет на глубину проплавления. Но эффект немного меньше по сравнению со скоростью хода дуги или сварочным током.
Методы сварки слева обычно приводят к более глубокому проплавлению по сравнению с методом сварки справа. Вы можете добиться максимального провара, используя сварку с обратной стороны и угол наклона горелки 25 градусов. Но больший угол наклона резака приводит к нестабильности дуги, что предотвращает более глубокое проникновение.
4. Скорость наплавки
Скорость наплавки зависит от скорости подачи проволоки. Вы можете изменить ток и расстояние от наконечника до металла для достижения определенной скорости наплавки.
Небольшое расстояние между наконечником и металлом приводит к низкой скорости подачи проволоки и высокому сварочному току. Результатом будет низкая скорость наплавки и более глубокое проникновение. Проникновение будет выше, поскольку при заданном токе осаждается меньше металла.
Для достижения более мелкого проникновения подходит большое расстояние от наконечника до металла.Это приведет к низкой скорости наплавки при заданном токе, что приведет к меньшему проплавлению сварного шва. Наплавленный металл шва с большим количеством наплавленного металла снижает силу дуги, что предотвращает более глубокое проплавление.
Каковы преимущества сварки с полным проплавлением?
Сварной шов с полным проплавлением имеет ряд преимуществ по сравнению со сварным швом с частичным или малым проплавлением.
1. Коррозионная стойкость
Металлы, сваренные швом с полным проплавлением, обладают более высокой устойчивостью к коррозии.При сварке с полным проплавлением образуется линия плавленного соединения. Напротив, линия стыка не заполнена частичным проникновением. Пространство между соединениями приводит к возникновению разности потенциалов, что приводит к коррозии металла.
2. Более прочный сварной шов
Как упоминалось ранее, сварной шов с полным проплавлением дает намного более прочный сварной шов по сравнению со сварным швом с частичным проплавлением. Шов заполняется сварным швом с полным проплавлением. Это приводит к увеличению объема металла, что приводит к повышению устойчивости к нагрузкам.
Напротив, сварные швы с методом частичного проплавления имеют зазоры на линии стыка. В промежутках сосредоточено геометрическое напряжение.
3. Полное слияние
Сварной шов с полным проплавлением охватывает большую площадь. Более широкое проникновение увеличит вероятность того, что наполнитель достигнет корня, что приведет к полному сплавлению. В случае частичного проплавления сварного шва и, как следствие, более узкого профиля проплавления, существует вероятность неполного плавления у основания металла.
Достижение полного сплавления у основания сварного шва имеет решающее значение.В случае, если расстояние от наконечника до металла велико, а электрод неправильно направлен на корень, отсутствие плавления может создать структурные проблемы получаемого металлического продукта.
Есть ли недостатки у сварки с проплавлением?
Хотя при полном проникновении металл становится более прочным, у этой техники есть определенные недостатки, которые вам следует знать.
1. Сварочная ванна
Первая проблема при сварке с полным проплавлением заключается в том, что в некоторых ситуациях это может привести к слишком большому количеству примесей в основном металле.Более высокая глубина проплавления приводит к увеличению объема основного металла. Увеличение объема основного металла может привести к образованию сварочной ванны, что увеличивает вероятность появления трещины.
Примеры, когда сварка с глубоким проплавлением не подходит, включают сварку металлов с более высоким содержанием свинца, фосфора или серы. Эти элементы имеют более низкие температуры плавления по сравнению со сталью. Это приводит к образованию сварочной ванны, которая снижает качество свариваемого металла. Лужа может перейти к центру сварного шва, что может привести к растрескиванию в центре.
2. Разбавление сварного шва
При наплавке и наплавке глубокое проплавление приводит к разбавлению сварочных отложений. Это приводит к более низкому профилю износостойкости свариваемого металла.
Наложение сварных швов с максимальным результатом проникновения в повышенной примесью базовой пластины и сварки металла. Противоположный случай — частичное проплавление, в результате чего металлические изделия становятся более качественными.
3. Прожигать
Сварка с глубоким проплавлением также не подходит для сварки металлических материалов.Слишком большой провар на тонком листе металла приведет к прожигу сварного шва металлического соединения.
Полное проплавление также не подходит, если необходимо выполнить корневой проход в стыке труб или открытых корневых стыках. Слишком большой провар может привести к прожиганию корня сварного шва, что приведет к снижению структурной целостности материала.
4. Растрескивание по средней линии
Наконец, глубокое проникновение также может привести к растрескиванию по средней линии. Чтобы избежать осевой линии или образования горячих трещин, необходимо соблюдать баланс между уровнем проникновения и шириной корневого прохода.Соотношение глубины проникновения и ширины не должно быть более 1: 1,2. Более высокое соотношение приведет к неправильной форме сварного шва, что приведет к снижению сопротивления нагрузкам.
Заключение
Полное проплавление подразумевает полное проплавление сварных швов. Следует правильно использовать сварные швы с полным проплавлением. Этот метод предпочтительнее частичного проплавления, чтобы обеспечить высокопрочный материал с большей устойчивостью к коррозии и нагрузкам.
Вы должны знать о минусах глубокого проплавления, таких как предрасположенность к прожогу, растрескиванию, образованию луж и в некоторых случаях разбавленным отложениям сварного шва.Информация в этом блоге поможет вам использовать правильные методы сварки для проекта.
Подобные сообщения:
Сопротивление корня трубных соединений из высокопрочного чугуна — UNITRACC
1 Введение
«Проникновение корней в канализацию и канализацию» признается препятствием для потока в контексте регулярных проверок внутренней части труб с помощью камеры. В частных водостоках проникновение корней может быть замечено только на поздней стадии, когда возникают засорения и обратное движение, а также связанные с этим последствия.В общественных канализационных системах проникновение корней является одной из наиболее частых причин повреждений [1]. Как в частных, так и в общественных канализационных системах эти участки трубопровода считаются не герметичными.
2 Рост корней и давление корней
Причины, по которым корни деревьев имеют тенденцию прорастать в стоки и канализацию, изучались в течение последних 15 лет. Важным результатом исследовательского проекта «Проникновение корней в канализацию и канализацию» [2] явилась разработка моделей, которые могут описывать проникновение корней в канализацию и трубопроводы.На основе результатов многочисленных раскопок были разработаны и описаны модель плотной ловушки и кислородная модель; они полезны для описания роста корней в подземном пространстве. В контексте дальнейшего исследовательского проекта [3] давление корней первичных корней и контактное давление вставных соединений были определены на различных трубных системах номинального размера DN 150, изготовленных из керамической глины, пластмасс и высокопрочного чугуна. Невозможно было сделать выбор между соединениями труб — например, в виде максимального зазора раструба — при котором можно ожидать минимального контактного давления, поскольку необходимо определить диапазон контактных давлений.Этот факт побудил Ассоциацию определить контактное давление в системе вставных соединений TYTON ® . С помощью следующих критериев, таких как
- ловушка плотности и кислородные модели,
- результаты исследования давления корней основных корней разных деревьев,
- исследования контактного давления на вставных трубных соединениях в контексте [3] и
- измерения на системе вставных соединений TYTON ® с максимальным зазором в раструбе
Удалось оценить сопротивление корня системы вставных соединений TYTON ® .
3 Плотностная ловушка модель
Вся окружающая среда зданий и их инфраструктура — это земное пространство, созданное антропогенным путем. В отличие от естественного грунта, он часто имеет более низкий уровень уплотнения и большие поровые пространства. Различия в плотности влияют на направление роста кончика корня. Эластичность калиптры (кончика корня) означает, что корни растут в направлении субстрата, в который легче всего проникнуть (, рисунки 1 и 2, ).Как правило, можно исключить рост корней обратно в зону с высоким уплотнением или с более плохими условиями проникновения. Корни «застревают» в участках почвы с большим потенциалом проникновения. Кольцевой зазор или пространство перед уплотнительным элементом также может, в зависимости от стыка труб, быть областью, к которой корни могут легко получить доступ. Они могут расти там несколько лет, прежде чем окончательно проникнут внутрь трубопровода. Для этого они должны преодолеть контактное давление герметика.
4 Кислородная модель
Наличие кислорода в почве имеет большое влияние на распространение корней.Все растительные организмы нуждаются в кислороде для поддержания своих метаболических процессов. Герметизация городских почв приводит к серьезному ограничению поступления кислорода в почву. Канализационные системы обычно работают как самотечные трубопроводы и в достаточной степени вентилируются через отверстия для технического обслуживания и осмотра (люки). Основная часть трубопровода заполнена воздухом. При использовании литых уплотнений в литейном материале могут возникать трещины из-за усадки. Таким образом кислород, содержащийся в воздухе, может попасть в окружающую среду труб и стыков труб в почве.При проектировании канализационных систем не учитывается газопроницаемость труб и стыков труб. Однако это также может повлиять на рост корней при недавно проложенных канализационных трубах. При использовании материалов для труб, которые не являются газонепроницаемыми, кислород может вытечь, даже если трубопроводы не повреждены. Согласно кислородной модели, корни растут к источнику кислорода и таким образом находят стык трубы.
Эта модель (окситропизм) подтверждается результатами [4]. На корнях проростков гороха (Pisum sativum L.) можно было продемонстрировать, что они следовали градиенту кислорода в направлении более высокой концентрации O2. Исходя из этого, целью исследовательского проекта «Проникновение корня в канализацию и канализацию — дополнительный проект» [3] было научно обосновать причины роста корня в трубопроводах и описать механизмы, лежащие в основе проникновения корня в трубопровод. и взаимодействие между свойствами трубы и проникновением в корень. Кроме того, должны были быть разработаны предложения по процедурам испытаний, чтобы реалистично проиллюстрировать механические и биологические процессы, вовлеченные в проникновение корня, чтобы соединения труб можно было оценить с точки зрения их сопротивления проникновению в корень.Это включало описание различий в поведении роста различных видов деревьев как характеристик разных корневых систем. Особое внимание уделялось взаимодействию корней с различными соединениями труб DN 150 и механическим испытаниям для изучения сопротивления корня. Результаты этих исследований кратко изложены в Разделе 9 «Расследования и результаты».
5 Практические примеры
Влияние порового пространства на рост корней можно увидеть на рисунках с 3 по 6 .Для испытаний в Ботаническом саду Рурского университета Бохума, показанных на Рисунках 3–5 , в сеялках было создано искусственное наслоение двух субстратов с разными уровнями пористости. Компост того типа, который используется для выращивания в контейнерах в Ботаническом саду Рурского университета Бохума, был выбран в качестве субстрата с высокой долей пор и хорошими характеристиками проникновения корней. Бентонит был выбран для воспроизведения участков почвы с низкой долей пор и плохими характеристиками проникновения корней.Используемый материал состоит из глинистых минералов со средним размером частиц 0,063 мм. Небольшой размер частиц приводит к тому, что необходимые для роста корней пространства (поры почвы) размером от 100 мкм и выше отсутствуют [5]. Подложки наносили попеременно горизонтальными слоями вокруг центральной колонны одной из подложек. Слоистую структуру можно увидеть в схематическом продольном разрезе, представленном на , рис. 3 . На рис. 4 показана сеялка, открытая с одной стороны.
Простой взгляд на это изображение дает намек на то, что корни не выросли в бентонит с малым размером пор, что было подтверждено после прорезания сеялки, включая почву, которая в нем содержалась, и промывки корней водой. Корни распространились через компост до дна сеялки ( Рис. 5 ). Между слоями бентонита они фактически доходили почти до вертикального края сеялки [2].
Это смоделированное в лаборатории поведение роста корней было подтверждено на практике.Корневая система, показанная на рисунке Рисунок 6 была найдена на глубине 7 м. Корни распространились по служебному трубопроводу к дому, подключенному через показанный фитинг к канализационной системе. Они продолжали прорастать в траншею для основной канализационной трубы и в пространство вокруг канализации. На этой глубине растущей корневой системе требуется достаточное количество кислорода. Есть основания полагать, что этот кислород поступал из негерметичных муфт соединительного трубопровода дома и основной канализации по линиям кислородной модели.
6 Характеристики различных корневых систем
Возможные эффекты различий в анатомической структуре корней были исследованы с помощью измерений корневого давления на первичные корни. В основном, у корней голосеменных (хвойных деревьев) было более низкое корневое давление, чем у корней покрытосеменных (широколиственные деревья). Пиковые значения корневого давления голосеменных растений варьировались от 4,0 бар для корней араукарии (Araucaria araucana) до 8.8 брусков для корней сосны (Pinus pinea). Давление корней покрытосеменных (широколиственных деревьев) также варьировалось. В качестве нижнего пикового значения было измерено 8,8 столбца для корней робинии, а в качестве верхнего предела было зафиксировано краткое значение 11,9 столбца для корней дуба. В среднем корневое давление составляет от 4 до 8 бар. Результаты измерений корневого давления сведены в Таблица 1 .
Давление корней в идеальных условиях для роста используемых корней саженцев — в каждой точке корни постоянно подавались вода и кислород [Источник: EADIPS / FGR]
| Виды | Измерение серия | Измерение период [ч] | Максимальное давление [бар] | Средние значения [бар] |
|---|---|---|---|---|
| Pisum sativum L./ Горох | 1 | 62,5 | 4,9 | 4,07 |
| 2 | 62,5 | 5,9 | ||
| 3 | 62,5 | 2,5 | ||
| Quercus robus L. / Дуб обыкновенный | 1 | 50,0 | 1,2 | 8.42 |
| 2 | 50,0 | 5,9 | ||
| 3 | 64,0 | 11,9 | ||
| 4 | 46,0 | 10,8 | ||
| 5 | 46,0 | 12,3 | ||
| Робиния псевдоакация L. / Недостаток саранчи | 1 | 58.0 | 8,8 | 6,43 |
| 2 | 58,5 | 8,4 | ||
| 3 | 48,0 | 6,7 | ||
| 4 | 48,0 | 6,5 | ||
| 5 | 25,0 | 3,7 | ||
| 6 | 22.0 | 4,5 | 6,28 | |
| Pinus pinea L. / Сосна / Итальянская каменная сосна | 1 | 700,0 | 3,6 | |
| 2 | 700,0 | 8,8 | ||
| 3 | 670,0 | 9,8 | ||
| 4 | 670.0 | 2,9 | ||
| Араукария араукана (МОЛИНА) К.КОЧ / Дерево-загадка обезьяны | 1 | 530,0 | 4,0 | 4,0 |
7 Соединения труб
Канализационные трубы чаще всего соединяются между собой с помощью вставных соединений с эластомером в качестве прокладки. По сравнению с другими системами они обладают тем преимуществом, что их сравнительно легко собрать даже в сложных условиях на стройплощадке.В последние годы разработка этих соединений была продвинута и оптимизирована с точки зрения конструктивных критериев. Наиболее часто используемым раструбным соединением для труб из высокопрочного чугуна является вставное соединение TYTON ® (, рис. 7, ). Он стандартизирован в диапазоне от DN 80 до DN 1400. С момента своего появления на немецком рынке в 1957 году он миллион раз зарекомендовал себя в трубопроводах питьевой воды, сырой воды и сточных вод. Основные размеры этого соединения определены в стандарте DIN 28603 [6] для номинальных размеров от DN 80 до DN 1400.Герметизирующую функцию вставного соединения TYTON ® выполняет профилированная прокладка, состоящая из смеси более мягкой резины (уплотняющая часть) и более твердой резины (удерживающая часть) [7].СТАНДАРТНОЕ вставное соединение ( Рис. 8 ) сравнимо с вставным соединением TYTON ® по своей конструкции и функциям. В Германии размеры его соединений определены в стандарте DIN 28603 [6] (форма C) от DN 80 до DN 2000. Прокладка состоит из резины с одной степенью твердости.
8 Взаимодействие между стыками труб и корнями
Для проверки соединений труб в лабораторных испытаниях моделируются ситуации нагружения, подобные тем, которые могут возникнуть в траншее для труб, чтобы гарантировать качество труб и соединений труб. Корни, которые врастают в стыки труб и приводят к утечкам, представляют собой еще не определенную нагрузку на стыки труб. Нагрузка, представленная ростом корня, была впервые описана в контексте [2]. В соответствии с этим корни не только проникают сквозь стыки труб, в которых отсутствует герметичность.Действительно, согласно общепринятым технологическим правилам, они могут преодолевать «устойчивые к корням» стыки труб, поэтому предположение, что «водонепроницаемые стыки труб означают герметичные стыки труб», применимо не во всех случаях. Результаты исследований в Швеции [8] и Австралии [9] подтверждают эти наблюдения. Стратегии, с помощью которых корни могут уничтожить стык труб, уже описаны в [2]. Особое внимание уделяется используемому материалу трубы, геометрической конструкции стыка трубы и применяемой эластомерной прокладке.Сумма свойств этих отдельных компонентов влияет на рост корней в области стыков труб. При этом необходимо учитывать дополнительные факторы, такие как снабжение корней кислородом через систему трубопроводов (диффузионная герметичность материалов труб и вставных соединений).9 Расследования и результаты
В контексте [3] контактные давления были определены в общей сложности на одиннадцати различных вставных соединениях номинального размера DN 150 и OD 160.Это включало три вставных соединения для труб из керамической глины, четыре для труб из ПВХ, три для труб из полипропилена и одно вставное соединение для труб из ковкого чугуна. Результаты испытаний контактным давлением на вставном соединении DN 150 TYTON ® без сдвигающей нагрузки и под действием сдвигающей нагрузки показаны на рис. 9 (все результаты приведены в таблице 18 в [3]. ). В качестве среднего контактного давления без воздействия сдвигающей нагрузки для системы вставных соединений TYTON ® было определено значение 22,2 бара.Под действием сдвигающей нагрузки среднее контактное давление 17,5 бар было создано на ненагруженной стороне системы вставных соединений TYTON ® .10 Риск проникновения и сопротивление корня в свете Технических правил
В соответствии с DIN 4060 [11] корневое сопротивление стыков труб считалось подтвержденным в Германии, если стыки труб под сдвигающей нагрузкой выдерживали испытания на герметичность под положительным и отрицательным давлением ( Рисунок 10 ). Это было сделано в предположении, что корни могут проникать только в негерметичные стыки труб.
Это было изменено на основе результатов исследований, описанных в [2] и [3], и было учтено при публикации междисциплинарных правил «Деревья, подземные трубопроводы и коллекторы», которые появляются с тем же текстом, что и DWA-M. 162 [12], техническое описание DVGW GW 125 и FGSV 939.
Важным изменением является то, что в двух главах «Герметичность и сопротивление корня» ([12], глава 5.5) и «Соединения труб» ([12], глава 5.6) впервые описывается риск проникновения для герметичных соединений труб ([12] , глава 5.5): «Корни могут вырасти не только в негерметичные трубы и соединения труб, но и в плотные соединения труб, которые не оказывают достаточного сопротивления корням».
Далее говорится: «При новых конструкциях и правильном изготовлении трубных соединений (например, в соответствии с DIN EN 1610 / DWA-A 139 для канализации) можно предположить, что опасность проникновения корня в трубопровод мала. Чтобы повысить сопротивление проникновению корней, могут быть приняты дополнительные конструкционные меры безопасности ([12], глава 7) ».
В качестве дополнительных конструктивных мер безопасности [12] в главе 7.2.2 описаны меры, которые принимаются непосредственно в районе подземных трубопроводов и траншей: так называемые пассивные защитные меры. Среди пассивных защитных мер:
- использование материала для заполнения малопористого пространства в траншее для труб и трубопроводов,
- установка обсадных труб (защитных труб) вокруг трубопровода,
- установка плит или защитных покрытий в траншею для труб,
- выбор корневых прочных соединений труб, другое оборудование.
Таким образом, из [12], глава 5.5 технического паспорта «Деревья, подземные трубопроводы и канализационные трубы» можно сделать вывод о риске проникновения в вставные соединения, даже если соединение труб установлено правильно. Вставное соединение с наименьшим риском проникновения или вообще без него будет классифицировано как устойчивое к корням в соответствии с [12], глава 7.
11 Измерение контактного давления на стыках труб
Геометрия герметика и результирующее влияние на создаваемое контактное давление не учитывались при испытании на сдвигающую нагрузку [11].Испытания показали только кратковременную нагрузку на герметик. Не обращали внимания на изменения свойств эластомера, такие как уменьшение контактного давления при более продолжительной сдвигающей нагрузке.
В стандарте EN 14741 [13] описан процесс испытаний для определения долговременных свойств уплотнения эластомерных прокладок путем экстраполяции и оценки давления уплотнения через 100 лет. В стандарте EN 14741 [13] процесс описывается следующим образом: «Давление уплотнения в стыке оценивается путем измерения давления, необходимого для подъема прокладки в каждой из трех трубок из ПТФЭ, которые равномерно распределены по окружности стыка между резиновую прокладку и внешнюю стенку гладкого конца или, если применимо, раструба ( Рисунок 11 ).В среде с регулируемой температурой и с увеличивающимися временными интервалами азот или воздух нагнетают через три гибкие трубки из ПТФЭ с постоянной скоростью потока 120 мл / мин. Измеряется давление p азота или воздуха, необходимое для достижения этой скорости потока. Давление p t измеряется в течение периода с увеличивающимися интервалами времени. Экстраполированные линии регрессии для p t используются для вычисления оценочных значений p x через 100 лет и p y через 24 часа.”
Трубка из ПТФЭ (политетрафторэтилена):
растянутая трубка, которая обычно используется как термоусадочная трубка. Исходный диаметр и первоначальная толщина стенки после усадки определяются обычным образом. Следует отметить, что размеры в растянутом состоянии обычно не определяются. Определенную толщину стенки и определенный диаметр необходимо тщательно проверить. Указанные предельные отклонения следует рассматривать как руководство для поставщиков.
п т :
давление, измеренное [бар] в трубке из ПТФЭ при скорости потока 120 мл / мин в течение времени t [час]
п x :
экстраполированное давление через 100 лет [бар]
п y :
расчетное давление через 24 часа [бар]
Схема испытаний показана на Рис. 11 .Исследования проводятся в соответствии с EN 14741 [13] без влияния сдвигающих нагрузок. На рисунке 12 показана типичная процедура измерения контактного давления и последующей экстраполяции в качестве примера.
12 Подтверждение устойчивости корней вставных соединений TYTON
®В приведенном ниже испытании показано, как можно продемонстрировать сопротивление корня вставных соединений на основе стандарта EN 14741 [13] с помощью модифицированного испытания для определения долговременных свойств уплотнения эластомерных прокладок путем оценки давления уплотнения.Для применения и последующей интерпретации результатов должны быть соблюдены следующие условия:
- Материалы труб и вставные соединения, использованные в контексте испытаний, являются очевидными диффузионно-непроницаемыми [15], так что нет подачи кислорода к корням траншеи трубопровода через систему труб.
- Для испытываемого вставного соединения определяются минимально возможные контактные давления. Для этого необходимо соблюдать граничные условия для типовых испытаний трубных соединений из чугуна с шаровидным графитом.Используются вставные соединения с максимальным кольцевым зазором в соответствии с EN 598 [10], нагруженные поперечной нагрузкой в выровненном положении.
- Даже толщина стенки используемого средства измерения (трубки из ПТФЭ) может влиять на измерение контактного давления. Чтобы принять это во внимание, перед испытанием диаметр гладкого конца должен быть уменьшен на соответствующую величину (например, путем «точения»).
- Испытания проводятся типично для групп DN, описанных в EN 598 [10], на вставных соединениях DN 200, DN 400 и DN 800.
Схема испытаний показана на рис. 13 , .
Несмотря на спецификации EN 14741 [13], четыре, а не три трубки из ПТФЭ расположены на ненагруженной стороне соосно установленного и нагруженного поперечным усилием соединения, представленного на Рис. 13 между уплотнением и поверхностью гладкий конец согласно Рисунок 14 в положениях 0 °, 45 °, 90 ° и 300 °. После этого давление, необходимое для подъема уплотнения, измеряется в соответствии с EN 14741 [13] через указанные интервалы.
Сопротивление корня вставного соединения, прошедшего испытания, считается доказанным, если контактное давление между эластомерным уплотнением и гладким концом вставного соединения больше, чем среднее давление кончиков корня, и, следовательно, ширина уплотнительной поверхности может также считается достаточно большим, чтобы вырезать кончики корней из кислорода, доступного в поровом пространстве почвы (диффузионная герметичность).Это относится к системе вставных соединений TYTON ® и СТАНДАРТНОЙ системе, если давление через 100 лет, экстраполированное в соответствии с описанным здесь процессом на основе EN 14741 [13], определено для вставных соединений с максимальным зазор стыка согласно EN 598 [10] в среднем больше 7.0 баров.
С помощью описанного здесь процесса были проведены испытания для оценки давления уплотнения на вставных соединениях системы TYTON ® , номинальные размеры DN 200 и DN 400 [16]. Реальная испытательная установка показана на рисунках 15 и 16 .
Для вставного соединения TYTON ® DN 200 было оценено среднее значение 7,67 бар через 100 лет, а для системы вставного соединения DN 400 расчетное среднее значение составило 7,76 бар через 100 лет. В результате испытанные вставные соединения были классифицированы как устойчивые к корням.
Подробные сведения об измерениях, результатах и оценке включены в отчет об испытаниях № G 32 980 от 14.02.2013 — Определение долговременной герметичности вставных соединений TYTON ® с эластомерными прокладками путем оценки давления уплотнения на основе EN 14741 [13] — Iro GmbH Oldenburg [16]. Его можно просмотреть и / или загрузить на сайте www.eadips.org/gutachten/.
13 Резюме и прогноз
Причины проникновения корней деревьев в канализационные и канализационные трубы исследовались последние 15 лет.В качестве значимых результатов следует выделить следующие:
- модель ловушки плотности,
- кислородная модель,
- возможность проникновения корней в водонепроницаемые швы,
- влияние конструкции стыка и геометрии прокладки,
- диффузионная герметичность материала трубы и прокладки,
- последствия для Технических правил.
Системы труб из высокопрочного чугуна, включая их вставные соединения, очевидно непроницаемы для диффузии, что означает, что подача кислорода к корневой системе в траншеи трубы может быть исключена, и, следовательно, отсутствует значительный стимул для роста корней.Кроме того, в контексте [2] было показано, что контактные давления и контактные поверхности давления трубных соединений из высокопрочного чугуна намного выше среднего корневого давления, которое также было определено экспериментально.
В стандартах на изделия из чугуна с шаровидным графитом [10] обычные функциональные испытания на герметичность подвижных соединений труб проводятся с максимальным кольцевым зазором соединения при одновременном действии сдвигающей нагрузки. В новой редакции стандарта на продукцию EN 598 [10] требование и проверка устойчивости корней включены в стандарт в качестве дополнительного элемента в форме долгосрочного испытания.
На основании этих результатов можно сделать вывод, что в подземных помещениях требуется вставное соединение, устойчивое к корням. Одно из таких приложений описано в статье «Принцип губчатого города — от систем« труба-грунт »до систем« грунт-трубы »- решения с системами труб из высокопрочного чугуна».
Литература[1] Stein, D. и Kaufmann, O .: Анализ повреждений бетонных и керамических труб канализационных систем в Федеративной Республике Германия — West Korrespondenz Abwasser, 1993-02
[2] Штютцель, Т.и др .: Проникновение корней в канализационные системы и стоки, Рурский университет в Бохуме в сотрудничестве с IKT — Institut für Unterirdische Infrastruktur gGmbH, Отчет об исследовании июль 2004 г.
Скачать: www.ikt.de/website/down/f0108langbericht.pdf
[3] Stützel, T. u. a .: Проникновение корней в канализационные системы и стоки — Дополнительный проект, Рурский университет в Бохуме в сотрудничестве с IKT — Institut für Unterirdische Infrastruktur gGmbH, Заключительный отчет об исследовании, июнь 2007 г. Скачать: www.ikt.de/website/down/ f0160langbericht.pdf
[4] Портерфилд, Д. М. и Масгрейв, М. Е. (1998): Тропическая реакция корней растений на кислород — окситропизм у Pisum sativum L. Planta 206 (1): цитируется в Streckenbach, M .: Взаимодействие между корнями и технической инфраструктурой под землей. — Принципы и стратегии избежания проблем, Диссертация 2009-06, Загрузить: www.streckenbach.org/projekt_jul_2009.html
[5] Кунтце, Х., Рошманн, Г. и Швертфегер, Г.: Почвоведение, Thieme-Verlag, 5-е издание 1994
[6] DIN 28603: 2002-05
[7] Электронная книга 10.2015, глава 8, Загрузить: www.eadips.org/e-book-e/
[8] Ridger, D. u. a .: Оценка испытаний бетонных и ПВХ труб, Опубликовано в: Заключительный научный отчет COST Action C 15 «Улучшение отношений между технической инфраструктурой и растительностью», 2008 г.
[9] Уиттл, А .: Сопротивление стыков труб с эластомерным уплотнением к проникновению в корни деревьевm, 2003-07 гг.
[10] EN 598: 2009-10
[11] DIN 4060: 1998-02
[12] DWA-M 162: 2013-02
[13] EN 14741: 2006-05
[14] Scharwächter, D.: Длительная герметичность герметичных стыков в системах пластиковых труб без давления, Plastic Pipes XI, 2001
[15] Wolf, W .: fgr volume 10 (1975), p.