Пример регламента по работе с дефектами на проекте для статьи Alien bugs или пришельцы из мира дефектов, Павел Новик, ЗАО «Технологии качества», бренд A1QA I.Описание полей, принципы заполненияЗаводя дефект, заполняется следующие 8 полей:
II.Подробнее про заполнение каждого из них:Приоритет – это важность дефекта. До определенной степени величина субъективная, но есть ряд критериев, на которые можно опираться. Основными определяющими свойствами бага являются: значимость нерабочей функциональности, влияние дефекта, специфичность окружения и воспроизведения.
Тема – краткое (в пределах 20 слов) описание ошибки, состоит из трех частей: < где найден дефект>: <короткое описание самой ошибки и места проявления><влияние (импакт) на пользователя>
Описание – самое большое поле, предназначенное для подробного описания проблемы. Состоит из следующих частей:
Компоненты – указывается модуль, для которого актуален дефект. Список доступных значений: Модуль регистрации; Модуль поиска; Модуль настроек, Общее; Проявляется в версиях – указывается версия билда, в котором был найден дефект. Окружение – указывается устройство и версия iOS (опционально несколько), на которых воспроизводится дефект. Исправить в версиях – указывается версия билда, в котором должен быть исправлен дефект. Вложение – логи, скриншоты или видео при необходимости, которые помогут понять путь воспроизведения дефекта и в чём его суть. Необходимо прикладывать для всех заводимых дефектов (для всех функциональных дефектов обязательно необходимо прикладывать логи). III.Общие правила заведения дефектов:Перед тем, как регистрировать ошибку в баг-трекер, необходимо провести как минимум три основных действия:
IV.Описание типичного жизненного цикла дефектов.
Возможные состояния и резолюции описаны в Jira, a также дополнительно приведены на скриншоте ниже. V.Пример описания дефекта несоответствующего регламенту.{необходимо привести описание реального дефекта с вашего проекта} VI.Пример описания дефекта соответствующего регламенту.{необходимо привести описание реального дефекта с вашего проекта} |

 п.
п. Для удобства восприятия шаги нумеруются: 1…2…3… В описании шагов важно найти тонкую грань между избыточностью (описание каждого движения мышкой) и недостаточностью (пропуск важных для воспроизведения шагов или отсутствие описания, где находится то окошко, в котором воспроизвелась ошибка).
Для удобства восприятия шаги нумеруются: 1…2…3… В описании шагов важно найти тонкую грань между избыточностью (описание каждого движения мышкой) и недостаточностью (пропуск важных для воспроизведения шагов или отсутствие описания, где находится то окошко, в котором воспроизвелась ошибка).
| Вид проявления дефекта | Наименование дефекта | Причины возникновения дефекта |
| Покрытия из рулонных материалов | ||
| Протечки, сырость на потолках и стенах; ухудшение температурно-влажностного режима в помещениях; разрушение отделочных покрытий |
Трещины в водоизоляционном слое и пароизоляции | Механические воздействия на рулонный ковер при его устройстве. Усушка или разбухание плит утеплителя. Деформации несущих конструкций и основания. Образование льда на покрытии. Сплошное приклеивание первого слоя к основанию Усушка или разбухание плит утеплителя. Деформации несущих конструкций и основания. Образование льда на покрытии. Сплошное приклеивание первого слоя к основанию |
| Пробоины в водоизоляционном слое | Механическое воздействие на рулонный ковер | |
| Влажность утеплителя превышает нормативную величину | Применение влажного утеплителя или увлажнение его во время устройства покрытия. Наличие непроклеенных участков в стыках полотнищ. Неплотности в местах примыкания рулонного ковра к стенам, парапетам, стоякам, шахтам. Отсутствие пароизоляции в совмещенном покрытии. Воздушная прослойка слабо вентилируется | |
| Трещины и отслоения в местах примыкания водоизоляционного слоя к выступающим конструкциям | Перегиб рулонного материала под углом 90° вместо 135°. Некачественное приклеивание полотнищ в местах изгиба. Механическое воздействие на ковер в период эксплуатации здания. Отсутствие выкружек. Не оштукатурены кирпичные поверхности в местах наклейки рулонного материала | |
Промерзание покрытия. Ухудшение температурно-влажностного режима в помещениях. Разрушение отделочных покрытий Ухудшение температурно-влажностного режима в помещениях. Разрушение отделочных покрытий |
Влажность утеплителя превышает нормативную величину | Проникание влаги через трещины, пробоины и неплотности в рулонном ковре и примыканиях. Выпадение конденсата внутри утеплителя в результате проникания влажного воздуха из помещения. Воздушные прослойки и каналы не вентилируются. Патрубок водоприемной воронки не утеплен |
| Скопление воды на покрытии. Возрастание нагрузки на покрытие | Отсутствуют уклоны в водоизоляционном слое | Брак при устройстве теплоизоляции и выравнивающей стяжки. Водоприемные воронки установлены на повышенных участках |
| Прогибы в водоизоляционном слое (блюдца) | Некачественное выполнение стяжки. Просадка утеплителя и стяжки. Прогиб несущих конструкций | |
| Водоприемные воронки забиты мусором | Отсутствие должного ухода за кровельным покрытием | |
| Сечение выходного отверстия у водоприемной воронки недостаточное | Ошибка проектирования. Установка водоприемной воронки с меньшим сечением выходного отверстия Установка водоприемной воронки с меньшим сечением выходного отверстия |
|
| Предаварийные неисправности. Возникновение опасности ухудшения эксплуатационных качеств покрытия | Разрушение защитного слоя | Применение остывшей мастики. Использование загрязненного гравия или крупного песка |
| Старение рулонного материала | Применение рулонного материала на основе картона. Слабая пропитка основы вяжущим. Воздействие ультрафиолетового излучения | |
| Старение мастики | Воздействие ультрафиолетового излучения | |
| Трещины в слое мастики | Деформации в основании. Температурные воздействия | |
| Вздутия (пузыри) на поверхности рулонного ковра | Отрыв верхнего слоя от нижележащих под воздействием давления водяных паров. Отрыв всех слоев рулонного ковра при сильном ветре. Отсутствие вентиляционной прослойки для выравнивания парциального давления водяных паров. Рулонные материалы были наклеены на влажное основание или во время дождя Рулонные материалы были наклеены на влажное основание или во время дождя |
|
| Водоизоляционный ковер заведен за парапет, стену, стояк, шахту на высоту менее 250 мм | Брак при выполнении работ | |
| Количество полотнищ, заводимых на стену, парапет, стояк, шахту, меньше проектного | Брак при выполнении работ | |
| Отсутствие штрабы, защитного пояска или уступов для предохранения затекания воды | Некачественное закрепление краев ковра и отсутствие герметизации шва | |
| Наличие неровностей в сопряжении рулонного ковра с металлическим сливом на свесе | Брак при выполнении работ | |
| В деформационном шве отсутствует компенсатор | Брак при выполнении работ. Нарушение проекта | |
| Нахлестка полотнищ в стыках меньше нормативной величины | Брак при выполнении работ | |
| Водоприемные воронки, трубостоки размещены на расстоянии менее 200 мм от стен, парапета, шахт, стояков | Ошибка проектировщика. Брак при выполнении работ Брак при выполнении работ |
|
| Снижение прочности утеплителя | Старение материала. Увлажнение | |
| Снижение прочности выравнивающей стяжки | Брак при выполнении работ. Применение раствора заниженной марки | |
| Разрушение сливов на парапетах, стенах, свесах | Коррозия металла. Механическое воздействие при сколе льда. Некачественное закрепление к основанию | |
| Покрытия из асбестоцементных и волнистых листов | ||
| Протечка, сырость на потолках и стенах. Ухудшение температурно-влажностного режима в помещениях. Разрушение отделочных покрытий | Трещины в листах | Механическое воздействие на лист. Воздействия низких температур, сильного ветра, льда |
| Пробоины (дыры) в листах | Старение материала. Механические воздействия. Окрашивание материала в местах забивки гвоздей | |
| Отсутствие листов или их части в кровельном покрытии | Срыв листов сильным ветром и воздействием отсоса.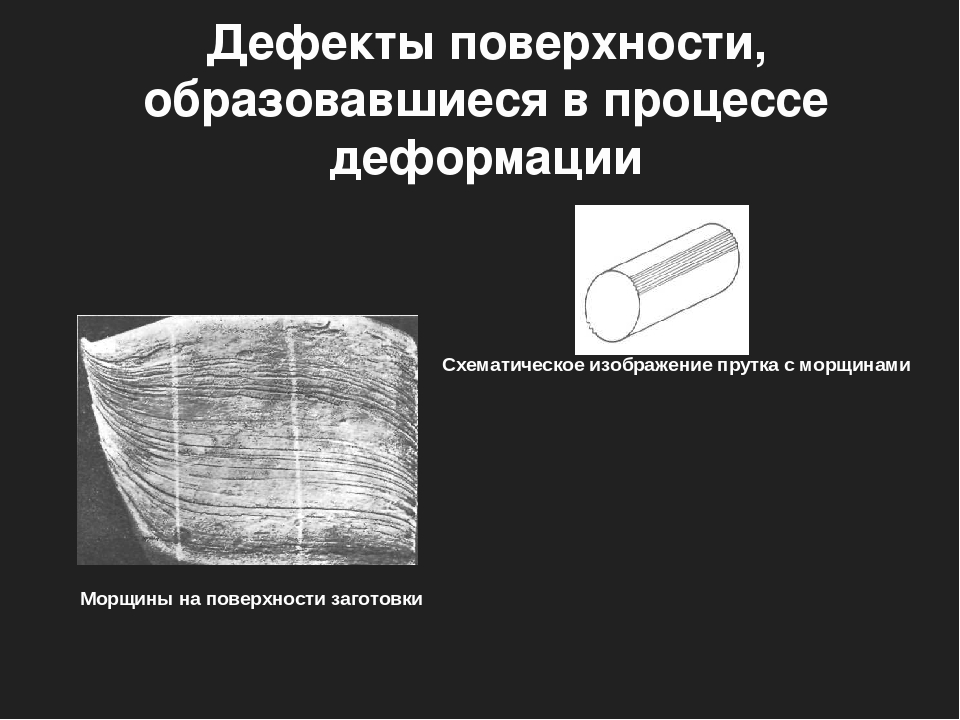 Смещение листов из ряда по причине некачественного закрепления Смещение листов из ряда по причине некачественного закрепления |
|
| Неплотности в местах соединения листов | Листы уложены без обрезки углов и без смещения на одну волну. Листы размещены в рядах с перекосом. Не закреплена на свесе уравнительная рейка. Листы имеют недопустимую величину искривления | |
| Неплотности в коньке | Детали конька уложены с перекосом. Не выполнена промазка швов | |
| Неплотности в местах примыкания к парапету, стене, трубе, фонарю, слуховому окну | Детали (уголки) примыкания не заведены в штрабу или под выступ. Штраба и борозда не заделаны раствором. Воротник неплотно прилегает к трубе, стояку | |
| Неплотности в разжелобках | Открытые торцы листов не промазаны раствором. Напуск асбестоцементных листов на лоток разжелобка менее допустимой величины | |
| Предаварийные неисправности. Возникновение опасности ухудшения эксплуатационных качеств кровельного покрытия | Сколы и незначительные трещины на листах | Воздействие инструментом при удалении льда и снега. Деформации кровельного покрытия. Воздействие сильного ветра. Воздействие льда Деформации кровельного покрытия. Воздействие сильного ветра. Воздействие льда |
| Неплотности в местах соединения листов | Некачественное выполнение работ. Воздействие сильного ветра | |
| Неплотности в коньке, местах примыкания листов к парапету, стене, трубе, фонарю, слуховому окну, в разжелобках | Брак при выполнении работ. Воздействие сильного ветра | |
| Плохое закрепление листов к обрешетке | Брак при выполнении работ | |
| Покрытия из листовой стали | ||
| Протечка, сырость на потолках и стенах. Ухудшение температурно-влажностного режима в помещениях. Разрушение отделочных покрытий | Неплотности в фальцах | Недостаточное обжатие (уплотнение) отгибов в фальцах. Малый отгиб имеет высоту менее 20 мм. Прогибы по длинной стороне картины. Коррозионное разрушение металла в фальцах |
| Пробоины, дыры, свищи в картинах, настенном желобе, картине свеса | Воздействие инструментом при удалении снега и льда. Воздействие инструментом при производстве работ. Коррозионное разрушение металла в картинах Воздействие инструментом при производстве работ. Коррозионное разрушение металла в картинах |
|
| Отсутствие отдельных картин покрытия | Воздействие сильного ветра. Воздействие сил отсоса при сильном ветре. Отсутствие кляммеров. Уменьшенное количество кляммеров для закрепления картин. Неудовлетворительное закрепление кляммеров к обрешетке. Некачественные фальцы | |
| Разрушение лотка и водоприемные воронки | Воздействие инструментом при удалении снега и льда. Коррозионное разрушение металла в деталях кровельного покрытия. Закрепление лотка и водоприемной воронки выполнено с отступлением от проекта | |
| Картины в разжелобках соединены одинарным лежачим фальцем и не промазаны суриком | Некачественное выполнение работы | |
| Промерзание перекрытия над верхним этажом | Влажность утеплителя превышает допустимую величину | Протечка кровельного покрытия. Задувание снега на чердак и его таяние |
Намокание участков фасада. Разрушение отделочных покрытий и материала стены Разрушение отделочных покрытий и материала стены |
Сгиб вниз картины свеса | Воздействие инструментом при удалении снега и льда со свеса. Отсутствие костылей под картиной свеса. Увеличение по сравнению с проектом расстояния между костылями |
| Картины свеса не соединены лежачим фальцем | Брак при выполнении работ | |
| Пробоины, дыры и свищи в картине карнизного свеса | Воздействие инструментом при удалении снега и льда. Коррозионное разрушение металла в картинах. Воздействие инструментом при производстве работ | |
| Отсутствие картины карнизного свеса | Некачественное закрепление картины. Воздействие сильного ветра. Воздействие сползающей глыбы снега или льда | |
| Отсутствие звеньев водосточной трубы, задвижка звеньев | Некачественное закрепление звеньев водосточной трубы. Воздействие сползающего по трубе льда. Случайные механические воздействия на трубу | |
Предварительные неисправности. Возникновение опасности ухудшения эксплуатационных качеств кровельного покрытия Возникновение опасности ухудшения эксплуатационных качеств кровельного покрытия |
Незначительные неплотности в фальцах | Брак при выполнении работ. Воздействие снега и льда. Коррозионное воздействие |
| Незначительные свищи, уменьшение толщины металла в картинах | Коррозионное воздействие. Воздействие инструментом при удалении снега и льда | |
| Прогибы в картинах | Расстояние между элементами обрешетки больше проектной величины. Нагрузка от снега или льда превышает расчетную. Уменьшение толщины металла в картинах | |
| Смещение звеньев водосточных труб | Ослабление стремян стяжек и хомутов. Механические воздействия на трубу | |
| Изменение сечения водосточных труб | Механические воздействия на звенья труб | |
Микроинвазивное витреоретинальное хирургическое лечение при двойном прободном ранении глаза Текст научной статьи по специальности «Клиническая медицина»
4° ^tl практИЧЕСКАЯ МЕДИЦИНА
‘2 (94) апрель 2016 г. / том 1
/ том 1
УДК 617.7-001.4/-003.6-089 Д.Г. АРСЮТОВ12
Республиканская клиническая офтальмологическая больница, 428014, г. Чебоксары, ул. Ашмарина, д. 85 Чувашский государственный университет имени И.Н. Ульянова, 428034, г. Чебоксары, пр. Московский, д. 45
Микроинвазивное витреоретинальное хирургическое лечение при двойном прободном ранении глаза
Арсютов Дмитрий Геннадьевич — кандидат медицинских наук, доцент кафедры офтальмологии и отоларингологии, главный врач, тел. (8352) 51-22-02, e-mail: [email protected]
Представлены результаты хирургического лечения пациентов с тяжелыми двойными прободными ранениями глаза с инородными телами различного происхождения и различных размеров (до 2,1 см длиной) с применением микроинвазивой 25-27 Ga витреоретинальной хирургии и методики трансвитреально-диасклерального подхода к удалению инородных тел. В ходе интраоперационного пособия по обеспечению сохранения целостности глазного яблока применен метод аллосклеропластики для закрытия крупных зияющих дефектов в задней стенке глаза с последующей газо-воздушной тампонадой полости стекловидного тела. Представленная тактика хирургического лечения двойных прободных ранений глаза позволяет получить высокий функциональный результат и добиться сохранения нормальной анатомии глазного яблока.
В ходе интраоперационного пособия по обеспечению сохранения целостности глазного яблока применен метод аллосклеропластики для закрытия крупных зияющих дефектов в задней стенке глаза с последующей газо-воздушной тампонадой полости стекловидного тела. Представленная тактика хирургического лечения двойных прободных ранений глаза позволяет получить высокий функциональный результат и добиться сохранения нормальной анатомии глазного яблока.
Ключевые слова: двойное прободное ранение глаза, внутриглазное инородное тело, трансвитреально-диасклеральный метод удаления инородных тел, аллосклеропластика дефекта склеры, газо-воздушная тампонада.
D.G. ARSYUTOV12
1 Republican Clinical Ophthalmology Hospital, 85 Ashmarin St., Cheboksary, Russian Federatio, 428014 2Chuvash State University named after I.N. Ulyanov, 45 Moskovskiy Ave., Cheboksary, Russian Federation, 428034
Microinvasive vitrectomy for double perforated eye injury
Arsyutov D. G. — Cand. Med. Sc., Associate Professor of the Department of Ophthalmology and Otolaryngology, Chief Physician, tel. (8352) 51-22-02, e-mail: [email protected]
G. — Cand. Med. Sc., Associate Professor of the Department of Ophthalmology and Otolaryngology, Chief Physician, tel. (8352) 51-22-02, e-mail: [email protected]
In the article are presented the results of surgical treatment of patients with severe double perforated eye injuries with foreign bodies of different origin and different sizes (up to 2,1 cm in length) with the application of microinvasive 25-27 Ga vitreoretinal surgery and methods of transvitreal and diascleral approach to the removal of foreign bodies. In the intraoperative manual on the conservation of the integrity of the eyeball is applied the alloscleroplasty method for closing large gaping defects in the back of the eye, followed by air-gas tamponade of the vitreous cavity. The presented tactics of surgical treatment of double perforated eye injuries allows obtaining high functional outcome and preservation of normal anatomy of the eyeball.
Key words: double perforated eye injury, intraocular foreign body, transvitreal and diascleral approach to the removal of foreign bodies, alloscleroplasty of scleral defect, air-gas tamponade. 41
41
родного тела большого размера в заднем отрезке глаза, при которых создается сложность обработки выходного раневого отверстия [1, 2]. В этих случаях высок риск отслойки сетчатки, субатрофии глазного яблока, полной потери зрения.
На сегодняшний день разработка эффективных методов комбинированной хирургии с пластикой дефекта заднего полюса глазного яблока является особо актуальной [3].
Цель работы — изучить эффективность комбинированного диасклерально-интравитреального подхода к удалению инородного тела с применением методик микроинвазивной 25-27 Ga витреорети-нальной хирургии с аллосклеропластикой дефекта заднего полюса глаза при двойных прободных ранениях.
Материал и методы
С февраля по декабрь 2015 года в БУ «Республиканская клиническая офтальмологическая больница» (г. Чебоксары) под наблюдением находились 4 пациента с двойными прободными ранениями глаза и локализацией инородного тела в заднем отрезке глаза. В трех случаях входное раневое отверстие было локализовано в склере, в одном случае — в роговице. Во всех случаях выходное раневое отверстие находилось в заднем полюсе глаза вне макулярной зоны и диска зрительного нерва (рис. 1, 2). При поступлении была диагностирована следующая сопутствующая патология: травматическая катаракта — у 1 пациента, гемофтальм различной степени выраженности — у 3 пациентов. Острота зрения травмированного глаза составляла от 0,02 до 1,0. Возраст пациентов варьировал от 19 до 45 лет.
Чебоксары) под наблюдением находились 4 пациента с двойными прободными ранениями глаза и локализацией инородного тела в заднем отрезке глаза. В трех случаях входное раневое отверстие было локализовано в склере, в одном случае — в роговице. Во всех случаях выходное раневое отверстие находилось в заднем полюсе глаза вне макулярной зоны и диска зрительного нерва (рис. 1, 2). При поступлении была диагностирована следующая сопутствующая патология: травматическая катаракта — у 1 пациента, гемофтальм различной степени выраженности — у 3 пациентов. Острота зрения травмированного глаза составляла от 0,02 до 1,0. Возраст пациентов варьировал от 19 до 45 лет.
Наличие инородного тела было подтверждено офтальмоскопически, рентгенологически, с помощью ультразвукового исследования [4]. У троих паци-
ентов инородное тело имело металлическую природу, различные размеры — длиной от 0,2 до 2,1 см, шириной — 0,1-0,15 см. В одном случае инородное тело было представлено осколком стекла длиной 1 см, шириной 0,4 см, толщиной 0,3 см (рис. 1, 3, 4).
В одном случае инородное тело было представлено осколком стекла длиной 1 см, шириной 0,4 см, толщиной 0,3 см (рис. 1, 3, 4).
Методика хирургического лечения.
После проведения первичной хирургической обработки все пациенты были прооперированы на 2-7 сутки с применением микроинвазивных 25-27 Ga витреоретинальных технологий. Из-за выраженного помутнения хрусталика в одном случае первым этапом была выполнена факоэмульсификация катаракты с имплантацией иОл. Во всех случаях была проведена субтотальная витрэктомия. Ввиду больших размеров инородного тела, сложности их выведения без дополнительной травматизации структур глаза, был осуществлен комбинированный (трансвитреально-диасклеральный) подход к их удалению. У всех пациентов диагностировано зияющее выходное отверстие в заднем полюсе глазного яблока. Была проведена эндолазерокоагуляция сетчатки вокруг места дефекта задней стенки глаза перед извлечением инородного тела для профилактики отслойки сетчатки. Для пластики дефекта склеры использовался аллотрансплантат, заводимый субтеноново к месту дефекта склеры шпателем. В одном случае была предпринята попытка силиконовой тампонады полости стекловидного тела, но силиконовое масло эвакуировалось через дефект в задней стенке глаза под коньюнктиву и в ретробульбарное пространство. Операцию во всех случаях завершали газо-воздушной тампонадой и позиционированием пациента на 3-5 суток лицом вниз. В случае, где предпринималась попытка силиконовой тампонады, через 1 месяц после операции был проведен второй этап хирургии — фа-коэмульсификация катаракты и резекция интра- и субтеноновых силиконовых кист.
Для пластики дефекта склеры использовался аллотрансплантат, заводимый субтеноново к месту дефекта склеры шпателем. В одном случае была предпринята попытка силиконовой тампонады полости стекловидного тела, но силиконовое масло эвакуировалось через дефект в задней стенке глаза под коньюнктиву и в ретробульбарное пространство. Операцию во всех случаях завершали газо-воздушной тампонадой и позиционированием пациента на 3-5 суток лицом вниз. В случае, где предпринималась попытка силиконовой тампонады, через 1 месяц после операции был проведен второй этап хирургии — фа-коэмульсификация катаракты и резекция интра- и субтеноновых силиконовых кист.
Рисунок 1.
Амагнитное инородное тело в заднем полюсе глаза (стекло)
Рисунок 2.
Глазное дно (7-е сутки после удаления инородного тела при сквозном ранении)
л
ОФТАЛЬМОЛОГЕ
*
42
ПРАКТИЧЕСКАЯ МЕДИЦИНА
‘2 (94) апрель 2016 г. / том 1
/ том 1
Рисунок 3, 4. Извлеченное инородное тело (стекло)
Результаты
Анатомическая целостность структур глазного яблока, не поврежденных инородным телом, была сохранена у всех пациентов. Отслойка сетчатки и геморрагические осложнения в раннем послеоперационном периоде и при последующем наблюдении в срок до полугода не были диагностированы ни у одного пациента. У троих пациентов в раннем послеоперационном периоде была достигнута полная стабилизация внутриглазного давления (в пределах 18-23 мм рт. ст.), обусловленное отсутствием фильтрации через задний раневой дефект вследствие «сухого» контакта газо-воздуш-ного пузыря с сетчаткой заднего полюса глаза, остающегося стабильным в течение всего периода наблюдения. В одном случае наблюдалась гипотония (6-8 мм рт. ст) в течение 5 суток после операции с последующим восстановлением офталь-мотонуса. Зрительные функции были сохранены у всех пациентов: у двоих пациентов — в пределах 0,2-0,4, у одного — 0,6, у четвертого пациента — 1,0.
Зрительные функции были сохранены у всех пациентов: у двоих пациентов — в пределах 0,2-0,4, у одного — 0,6, у четвертого пациента — 1,0.
Выводы
В случае двойных прободных ранений глаза целесообразно осуществлять комбинированный транс-витреально-диасклеральный подход к удалению инородного тела с применением микроинвазивной 25-27 Ga витреоретинальной хирургии с аллоскле-ропластикой дефекта заднего полюса глаза и одномоментной тампонадой полости стекловидного тела газо-воздушной смесью, что значительно минимизирует риск осложнений в ходе операции, позволяет сохранить нормальную анатомию глаза и получить высокий функциональный результат.
ЛИТЕРАТУРА
1. Захаров В.Д. Витреоретинальная хирургия. — М.,2003. — С. 110-122.
2. Чарльз С. , Кальсада Х., Вуд Б. Микрохирургия стекловидного тела и сетчатки. Иллюстрированное руководство. — М.: МЕДпресс-информ, 2012. — С. 331-336.
, Кальсада Х., Вуд Б. Микрохирургия стекловидного тела и сетчатки. Иллюстрированное руководство. — М.: МЕДпресс-информ, 2012. — С. 331-336.
3. Зернова Л.А. Первичная хирургическая обработка склеральных ран методом аллосклеропластики эписклеральной стороной: автореферат. — М., 2010.
4. Андреев А.А. Разработка этапности и объема хирургического лечения пациентов с внутриглазными инородными телами пре- и постэкваториальной зон: автореферат. — М., 2012.
Как проявляется дефект | Название дефекта | Из-за чего возникает дефект |
Дефекты кровли из рулонных материалов | ||
Сырые стены и потолки, протечки, холод и сырость в помещении, порча отделочных покрытий | Трещины в слое водоизоляции | Дефекты несущих конструкций. Повреждения рулонного ковра Разбухание или усушка утеплителя. |
Повреждения, дыры в слое водоизоляции | Физические повреждения | |
Несоответствие влажности утеплителя стандартам (влажность превышает стандарт) | Отсутствует пароизоляция совмещенного покрытия. Плохо проклеенные участки в стыках полотнищ. | |
Отслоение водоизоляции в местах прилегания к выступающий частям | К указанному дефекту приводит В местах наклейки рулонного материала должны быть оштукатурены поверхности. Если кирпичные поверхности не оштукатурены, это приводит к дефекту наклейки рулонного материала. Отсутствие выкружек. (Выкружка — непрямолинейный вогнутый облом, | |
Покрытия промерзают. Холод и сырость в помещении. | Влажность утеплителя больше нормы | Проникновение влаги через трещины, пробоины и неплотности в рулонном ковре и примыканиях. Внутри утеплителя выпадает конденсат (причина – проникновение влажного воздуха в помещение). Отсутствует вентиляция воздушных прослоек. Не утеплен патрубок вентиляционной прослойки. |
Застой воды на кровельном покрытии ( в результате чего увеличивается давление на кровлю). | Нет уклонов в слое водоизоляции | Брак при устройстве теплоизоляции и выравнивающей стяжки. |
Перегибы в слое водоизоляции | Дефекты монтажа стяжек. | |
Водоприемные воронки забиты мусором | Плохой уход за кровлей | |
Недостаточное сечение выходного отверстия водоприемной воронки. | Ошибка в проекте. | |
Опасность ухудшения эксплуатационных качеств кровли | Дефекты, порча защитного слоя | Недостаточно разогретая мастика. Загрязнения гравия или песка |
Рулонный материал стареет | Рулонный материал сделан на основе картона. Плохая пропитка основы вяжущим составом. Солнечные лучи. | |
Мастика стареет | Воздействие прямых солнечных лучей | |
Появление трещин в слое мастики | Воздействие температурных колебаний атмосферы | |
Пузыри на поверхности рулонного покрытия | Отрыв верхнего слоя от нижних слоев в результате погодных воздействий. | |
Заведение водоизоляционного ковра за парапет, стену, стояк, шахту на неверную высоту (меньше 250 мм) | Ошибки при монтаже | |
Малое количество полотнищ, заводимых на стену, стояк, шахту, парапет. | Ошибки при монтаже | |
Отсутствие защитного пояска или уступов для предотвращения затекания воды | Неправильно закреплены края рулонного ковра, не загерметизирован шов | |
Неровно сопряжен рулонный ковер и металлический слив на свесе | Некачественный монтаж | |
Отсутствие компенсатора в деформационном шве | Некачественный монтаж, несоблюдение проектных требований | |
Нарушение нормативной величины нахлестки полотнищ | Некачественный монтаж | |
Неправильное размещение водоприемных воронок и трубостоков (менее 200 мм от парапета, стен, шахт и стояков) | Ошибка в проекте. | |
Потеря утеплителем своей прочности | Утеплитель портится от времени. Излишнее увлажнение | |
Выравнивающая стяжка теряет прочность | Некачественный монтаж и некачественный раствор. | |
Порча сливов | Коррозия металла. Неосторожная сколка льда с кровли. Плохое укрепление сливов | |
Дефекты кровли из волнистых и асбестоцементных листов | ||
Сырые стены и потолки, протечки, холод и сырость в помещении, порча отделочных покрытий | Трещины листов | Механические повреждения. Атмосферные воздействия – холод, ветер. |
Образование пробоин | Порча материала от времени. Физические повреждения. Покраска материала в местах забитых гвоздей | |
Недостаточно листов в покрытии | Срыв листов сильным ветром и воздействием отсоса. | |
Неплотно соединены листы | Листы уложены косо. Листы имеют неверную величину искривления. Нет без обрезки углов и смещения на одну волну. Нет закрепления уравнительной рейки на свесе. | |
Дефекты конька (Неплотности) | Детали конька смонтированы косо. Не сделано промазывание швов | |
Неплотное примыкание к парапетам, фонарям, слуховым окнам, трубам. | Уголки примыкания не заведены в штрабу или под выступ. Плохая заделка раствором. Неплотное прилегание воротника к стояку | |
Дефекты (неплотности) разжелобков | Открытые части торцов листов не промазаны раствором. Недостаточный напуск асбестоцементных листов на лоток разжелобка | |
Ухудшение состояния кровли и ее эксплуатационных качеств | Трещины на листах | Механические повреждения при удаление льда. |
Неплотное соединение листов | Ошибки при монтаже кровли. Сильный порывистый ветер | |
Неплотности в коньке, местах примыкания листов к парапету, стене, трубе, фонарю, слуховому окну, в разжелобках | Ошибки монтажа. Сильный ветер. | |
Плохо прикреплены листы к обрешетке | Ошибки монтажа. | |
Дефекты кровли из листовой стали | ||
Сырые стены и потолки, протечки, холод и сырость в помещении, порча отделочных покрытий | Неплотные фальцы | Ошибки в отгибах фальцев. Коррозия металла в фальцах |
Дыры, свищи, пробоины в картинах, настенном желобе, картине свеса ( КРОВЕЛЬНАЯ КАРТИНА -заготовка из одного или двух листов кровельной стали с четырьмя отгибами по сторонам) | Неосторожное удаление снега и льда. | |
Отсутствие части картин покрытия | Атмосферные явления (сильный ветер). Отсутствуют кляммеры или их количество недостаточно. Плохое укрепление кляммеров к обрешетке. Некачественно сделанные фальцы | |
Разрушен лоток и водоприемные воронки | Неосторожное удаление льда и снега. Ошибки в проекте. | |
Картины в разжелобках некачественно соединены фальцем, плохое промазывание суриком или его отсуствие | Некачественный монтаж | |
Промерзание перекрытия над самым верхним этажом | Утеплитель имеет влажность, превышающую нормы | Протечка кровли. Занос снега на чердаки. |
Отсыревание некоторых участков фасада. | Сгибание картины свеса | Повреждения кровли при неосторожном удалении снега и льда. Отсутствуют костыли под картиной свеса. Неверные расстояния между костылями (больше нормы) |
Отсутствует соединение лежачим фальцем картин свеса | Некачественное выполнение работ | |
Пробоины, дыры и свищи в картине карнизного свеса | Физическое неосторожное воздействие инструментом при удалении снега и льда. | |
Отсутствие картины карнизного свеса | Некачественное закрепление картины. Воздействие сильного ветра. Воздействие сползающей глыбы снега или льда | |
Отсутствие некоторых звеньев водосточной трубы | Некачественное закрепление звеньев водосточной трубы. Механические воздействия льда, движущегося по трубе вниз | |
Ухудшение качества кровли | Неплотные фальцы | Некачественно выполненные работы. Воздействия атмосферы. Коррозия металла. |
Незначительные свищи, уменьшение толщины металла в картинах | Воздействия атмосферы. Коррозия металла. Неосторожное удаление снега и льда | |
Картины прогибаются | Нарушение проектных норм в расстояниях между элементами обрешетки. Давление большого количества снега и льда. Недостаточная толщина металла в картинках. | |
Смещены звенья водосточной трубы | Ослаблены стремена хомутов и стяжек. Механические повреждения трубы | |
Нарушение сечения водосточной трубы | Физические повреждения | |


 Водоприемные воронки установлены на повышенных участках
Водоприемные воронки установлены на повышенных участках Отрыв всех слоев от ветра. Нет вентиляционной прослойки, служащей выравниванию парциального давления водяных паров. Наклеивание материала рулона
Отрыв всех слоев от ветра. Нет вентиляционной прослойки, служащей выравниванию парциального давления водяных паров. Наклеивание материала рулона Некачественный монтаж.
Некачественный монтаж. Смещение листов из ряда по причине некачественного закрепления
Смещение листов из ряда по причине некачественного закрепления Атмосферные воздействия.
Атмосферные воздействия. Коррозия (разрушение) металла в картинах
Коррозия (разрушение) металла в картинах Коррозия металла в картинах. Механические повреждения инструментом при производстве работ.
Коррозия металла в картинах. Механические повреждения инструментом при производстве работ.
КРЫЛЬЦО, ВЕРАНДА И ТЕРРАСА В ЧАСТНОМ ДОМОСТРОИТЕЛЬСТВЕ
В новом материале разбираемся, что такое крыльцо, веранда и терраса в жилых и садовых домах, и в чем их особенности
Эти конструктивные элементы носят разное назначение, отличаются площадью, типом фундамента и возможностью использования.
► КРЫЛЬЦО
Крыльцо — небольшая пристройка к основному зданию, обеспечивающая удобный вход в него.
Крыльцо выполняет утилитарную функцию – оно обеспечивает быстрый доступ к помещениям, расположенным внутри здания. Небольшая площадь и отсутствие стен делают его использование для иных нужд маловероятным.
Также крыльцо может быть использовано в качестве декоративного архитектурного решения и менять восприятие внешнего вида дома.
Обязательные элементы конструкции:
Для удобства и безопасности крыльцо часто оснащают перилами, а для защиты от атмосферных осадков – навесом. Кровля может быть заведена под общую крышу здания, а может выступать как отдельная конструкция.
Крыльцо возводится с использованием различных материалов и не связано фундаментом с основным зданием, что исключает повреждения и возможные дефекты.
Крыльцо – это узкоспециализированный элемент постройки, что отличает его от террасы или веранды, назначение которых гораздо шире.
Особенности:
— Не отапливается
— Небольшая площадь
— Отдельный фундамент и отсутствие стен
— Утилитарное назначение конструкции
! Крыльцо входит в общий контур здания, но при подсчете общей площади жилого помещения не учитывается
► ТЕРРАСА
Терраса – это дополнительная пристройка-настил, которая возводится на отдельном основании (обычно приподнятом на 15-45 см).
Особенности конструкции:
— отдельное основание или фундамент
— отсутствие стен и остекления
— может присутствовать навес, защищающий от осадков (фиксируется на прочных высоких столбах по периметру)
— могут присутствовать перила и ограждения, обеспечивающие безопасность
— вариативность конструкции: может устанавливаться с любой стороны дома, идти по периметру дома или стоять отдельно от основного здания и быть соединено с ним переходом
! Так как навес не является обязательным элементом, а утепление и отопление открытой площадки террасы не предусмотрено, может быть использована только в теплое время года.
! Терраса не входит в общую жилую площадь дома и для строительства разрешительная документация не требуется
Основное отличие террасы от крыльца в более широком спектре использования. За счет большей площади и отсутствии узкоутилитарного назначения на террасе можно отдыхать, установить мангал, джакузи и другие необходимые для отдыха элементы.
За счет большей площади и отсутствии узкоутилитарного назначения на террасе можно отдыхать, установить мангал, джакузи и другие необходимые для отдыха элементы.
► ВЕРАНДА
Веранда – крытая пристройка к дому, возводимая на общем или отдельном фундаменте.
Особенности конструкции:
— не отапливается
— имеет общую или заводимую под основное здание кровлю
— может возводиться на общем с основным зданием фундаменте
— наличие большого по площади остекления с 2-3х сторон
— примыкает непосредственно к основному зданию и является его частью
Веранда может быть использована и как летняя кухня, и как гостиная, столовая или спальня в теплое время года. Так как для постоянного проживания она не подходит из-за отсутствия отопления, веранда не относится к жилым помещениям.
! При выполнении кадастровых работ по подготовке технического плана для постановки объекта недвижимости на кадастровый учет ВЕРАНДЫ включаются и учитываются при подсчете площади жилого/садового дома.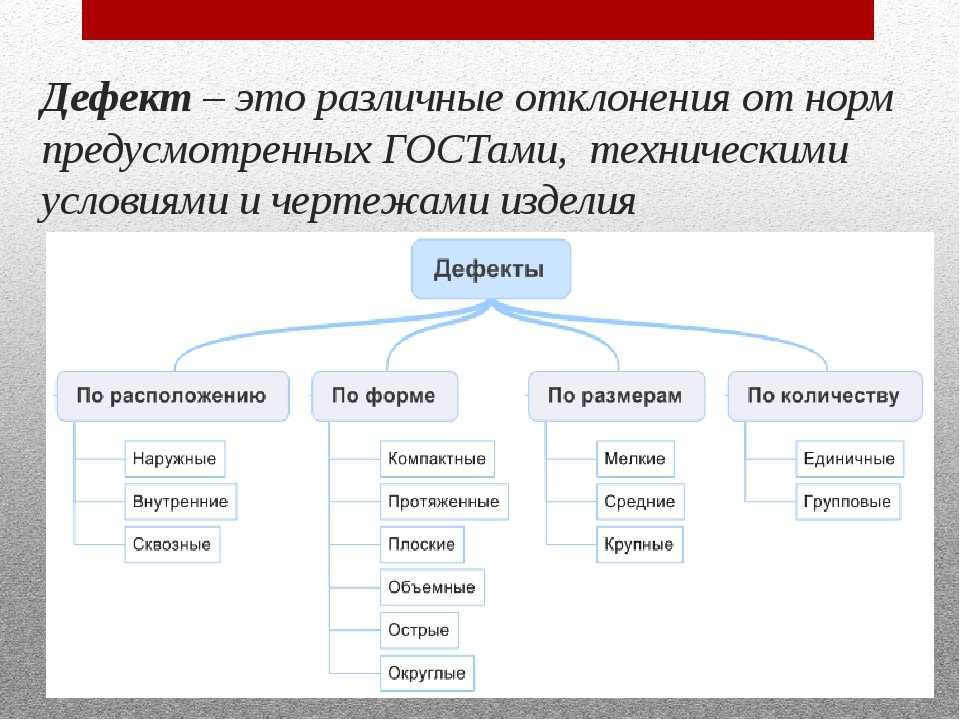
► РЕКОНСТРУКЦИЯ
Если веранда была пристроена к дому уже после постановки на кадастровый учет, то необходимо внести изменения о проведенной реконструкции в ЕГРН, так как изменились параметры дома, его площадь и иные характеристики.
Для этого потребуется вызвать кадастрового инженера для внесения изменений в технический план дома и дальнейшего подачи обновленного документа в Росреестр на внесение изменений в их данные.
Еще больше полезных материалов в телеграм-канале @mobti и в социальных сетях МОБТИ
Теги: МОБТИ, просто и понятно, недвижимость, ИЖС, СНТ, технический план, 2020
Заведение коммерческого линолеума на стену
Существует превеликое множество способов заведения напольного покрытия на стену. К ним относятся разнообразные запатентованные материалы, инструменты и специальные технологии, ну и конечно уникальные ноу-хау навыки каждого мастера.
К ним относятся разнообразные запатентованные материалы, инструменты и специальные технологии, ну и конечно уникальные ноу-хау навыки каждого мастера.
Выбор того или иного технологического решения задачи заведения напольного материала на стену всегда индивидуален. Поэтому для правильной оценки стоимости работ потребуется определится с методом, материалами и объемами работ. В первую очередь, необходимо знание следующих параметров:
1. Высота заводения напольного покрытия на стену.
2. Количество внешних и внутренних углов заводимого на стену напольного покрытия.
1. Ваши требования к радиусу кривизны при заведении покрытия на стену.
Хороший эстетический вид и надежность достигается при использовании угловых специальных профилей.
Для мармолеума применяются профили с большим радиусом кривизны, для коммерческих ПВХ покрытий радиус может быть меньше. Для бытового линолеум радиус заведения может быть совсем небольшим, таким что можно заводить без профиля.
При заведении покрытия без направляющего профиля необходимо обеспечить радиус кривизны не более 5мм. Далеко не каждое покрытие способно выдержать такой залом, даже при нагревании.
Отход от технологии заведения напольного покрытия рано или поздно приводит к образованию дыр. Например, когда медицинское оборудование или мебель придвигают вплотную к стенке и этим продавливает напольное покрытие в месте изгиба.
2. Ваши требования к верхней кромке заводимого на стену покрытия.
Покрытие имеет некоторую толщину (от 2 мм и более) и находясь на открытых взгляду стенах может не соответствовать Вашим представлениям об эстетике помещения. Наиболее красивый срез удается получать на гомогенных ПВХ покрытиях коммерческого класса. А вот открытая торцевая кромка коврового покрытия или натурального линолеума практически всегда будет выглядеть не очень красиво.
Наиболее популярным решением скрытия торцевой части напольного покрытия заведенного на стену является применение специальных кромочных профилей. Эти профили промышленно изготавливаются небольшими предприятиям из пластика (ПВХ) или металла (алюминиевые сплавы).
3. Ваши требования к использованию угловой фурнитуры:
Для оформления углов стен в местах примыкания к напольному покрытию можно использовать непосредственно материал напольного покрытия или же специальную угловую фурнитуру.
Угловая фурнитура изготавливается из пластика или металла и имеет, как правило, устойчивую геометрическую форму. Углы без фурнитуры, когда угол отделывается только материалом напольного покрытия, каждый угол будет в чем-то уникален.
Поэтому когда на небольшом участке стены сосредоточено несколько углов мы рекомендуем использовать специальную угловую фурнитуру.
Но есть и ограничения: Если стены не сходятся строго под прямым углом (90), то фурнитуру скорей всего не удастся установить красиво.
С учетом российской специфики строительства и ремонта применение угловой фурнитуры в Москве не очень распространено.
4. Ваши требования к исполнению угловых швов заводимого на стену покрытия
Наиболее распространенными решениями являются прямой и V-образный шов угла. Но могут быть и Ваши уникальные дизайнерские пожелания.
Другой важный момент: применять или нет сварку углового шва. Надежней и практичнее всего сваривать шов горячим способом. Но это дорогая и кропотливая работа. Другой вариант — применение для заделки угловых швов декоративных шпаклевок или герметиков.
5. Ваши требования к бордюрному шву
Примыкание напольного покрытия к стене можно классифицировать по трем общим методам.
1. Когда покрытие с пола сразу, без лишних швов, накатывается на стену. Этот способ чаще применяют для небольших помещений, соизмеримых по стене с шириной рулона.
2. Когда напольное покрытие приклеивается отдельно и потом к нему плотно прирезается лента материала заводимого на стену. Этот способ более технологичен и позволяет достичь наивысшего качества отделки стен и углов.
3. Установка плинтуса — самый простой и наиболее распространенный способ.Заведение линолеума на стену является стандартным гигиеническим требованием для напольных покрытий лечебных и детских образовательных учреждений
Пол с покрытием, заведенным на стену, легче моется и грязь не скапливается в углах.
В технологическом плане заведение линолеума на стену требует предварительной установки пластикового или резинового профиля-плинтуса по которому затем покрытие накатывается на стену. При работе с натуральным линолеумом требуются специальные навыки, позволяющие избежать заломов материала.
Обучающее и рекламное видео, прочие информационные материалы из отрытых источников по теме сварке ПВХ
Общие требования к организациям, осуществляющим медицинскую деятельность: 4.3. Покрытие пола должно плотно прилегать к основанию. Сопряжение стен и полов должно иметь закругленное сечение, стыки должны быть герметичными. При использовании линолеумных покрытий края линолеума у стен могут быть подведены под плинтуса или возведены на стены. Швы, примыкающих друг к другу листов линолеума должны быть пропаяны. В вестибюлях полы должны быть устойчивы к механическому воздействию (мраморная крошка, мрамор, мозаичные полы и другие). Полы в операционных, наркозных, родовых и других аналогичных помещениях должны быть антистатическими. Полы в вентиляционных камерах должны иметь непылеобразующее покрытие.
Санитарно-гигиенические требования к стоматологическим медицинским организациям: 3.8. При отделке стоматологических кабинетов, в которых применяется ртутная амальгама: — полы должны настилаться рулонным материалом, все швы свариваются, плинтус должен плотно прилегать к стенам и полу;
Санитарно-эпидемиологические требования к устройству, оборудованию и эксплуатации фельдшерско-акушерских пунктов, амбулаторий: 4.4. Покрытия пола не должны иметь дефектов, должны быть гладкими, плотно пригнанными к основанию. При использовании линолеумных покрытий края линолеума у стен должны быть заведены на стены, или подведены под плинтуса или должны быть плотно закреплены между стеной и полом. Швы примыкающих друг к другу листов линолеума должны быть пропаяны.
Автоматизация тестирования вебсайта: Pytest, Allure, Selenium + несколько секретных ингредиентов
Уже больше года у Acronis новый логотип, о том, как он создавался можно прочитать тут. В рамках ребрендинга обновился и сайт www.acronis.com. Для построения сайта мы выбрали CMS Drupal. Сайт получился классный снаружи и непростой внутри: 28 локалей (!), отзывчивый веб-дизайн, карусели, визарды, всплывающие окна… в итоге пришлось задействовать множество различных модулей друпала, некоторые из них кастомизировать и писать собственные. При работе такой динамичной, сложной системы неизбежны ошибки. Их предотвращением и выявлением занимается команда контроля и обеспечения качества вебсайта. В этой статье мы хотим поделиться своим успешным рецептом автоматизации тестирования нашего сайта.
Почему и зачем?
К сожалению, на начальном этапе жизни нашего сайта на страже качества были, в основном, только возможности ручного тестирования. Учитывая короткие релиз-циклы, регрессионные тесты приходилось прогонять часто. Большое количество локалей усугубляло ситуацию (проверки умножались на количество локалей). Затраты на ручное тестирование были большими, а выгода автоматизации тестирования сайта очевидной — надо было действовать.
Чего хотим?
Итак, начинаем составлять требования к системе автоматического тестирования. Мы заметили, что на обработку достаточно простых дефектов тратится драгоценное время, захотелось минимизировать количество заводимых в баг-трекер дефектов, но не теряя такие ошибки из поля зрения. Поэтому работу с такими дефектами было решено возложить на систему отчетов автотестов. Требовалось, чтобы репортинг был очень удобным, понятным, информативным и приятным. Таким, чтобы любой член нашей дружной web-команды мог посмотреть отчет, быстро понять, в чем дело, и пофиксить дефект. Соответственно все смотрят: разработчики фиксят сайт, тестировщики фиксят тесты. В итоге имеем «зеленый сигнал светофора» и GO в продакшн.
Исторически сложилось ещё одно требование — язык разработки Python. Опуская подробности остальных пожеланий, мы получили перечень требований для будущей системы автоматического тестирования:
- Удобный, понятный, информативный репортинг;.
- Параллельный запуск тестов;
- Параметризация тестов;
- Наследования тестовых данных по локалям;
- Тестирование в среде, наиболее приближенной к пользователю;
- Минимизация ложных срабатываний;
- Удобная поддержка и доставка окружения для тестирования;
- Минимум собственной разработки и кастомизации;
- Open source;
- Python.
Муки выбора
Первым делом мы обратили внимание на Robot Framework. Это популярное решение, которое уже упоминалось в публикациях Acronis. Установили, начали пробовать. Плюсов, конечно, много, но сейчас не о них. Основное противопоказание для нас — это работа RF с параметризованными тестами и штатный репортинг результатов для них. Следует заметить, что абсолютное большинство задуманных нами тестов — это именно параметризованные тесты. В отчете RF, если один из случаев в параметризованном тесте провален, то весь тест помечается как проваленный. Также репортинг RF из коробки был неудобен для большинства наших ребят — много времени уходило на разбор результатов пробных автотестов. Своеобразное «простое» написание тестов при помощи ключевых слов в случает тестов посложнее оказалось непростой задачей. В то же время, благодаря в том числе и Хабру, мы обратили внимание на Allure от Яндекс.
Посмотрели, всем понравилось, у всех возникло желание работать с этой системой, что, надо заметить, очень важно. Мы изучили инструмент, убедились, что проект активно развивается, имеет много готовых адаптеров для различных популярных тестовых фреймворков, в том числе и для python. К сожалению (а может, и к счастью) оказалось, что для python существует адаптер только для фреймворка pytest. Посмотрели, почитали, попробовали — оказалось, крутая штука. Pytest много всего умеет, прост в использовании, расширяемый функционал за счёт готовых и собственных плагинов, большое интернет-сообщество, ну, а параметризация тестов на нем такая, как нам нужна! Запустили, написали пару пробных тестов, все отлично. Теперь дело за параллельным выполнением тестов, тут выбор решений невелик — только pytest-xdist. Установили, запустили и…
Ложка дегтя
Оказалось, что pytest-xdist конфликтует с Allure Pytest Adaptor. Начали разбираться, проблема известная, уже длительное время обсуждается и не решена. Также готового решения для наследовании тестовых данных по локалям найти не удалось,
Секретные ингредиенты
Чтобы обойти эти проблемы, мы решаем написать тул (обертку) на python. Эта обертка будет подготавливать тестовые данные с учетом наследования, передавать их тестам, передавать тестам данные тестовой среды (браузер, например), а также запускать тесты в заданное количество потоков. После выполнения тестов — объединять полученные в разных потоках отчеты в один и публиковать конечные данные на вебсайт.
Параллелинг решили реализовать достаточно просто, как параллельный вызов каждого единичного теста через командную строку. При таком подходе пришлось реализовывать передачу тестовых данных в тест самостоятельно. Благодаря фикстурам в pytest это дело нескольких строк. Важно! Также необходимо закомментировать пару строк в (allure-python/allure/common.py), отвечающих за удаление «старых» файлов в директории отчетов allure адаптера.
Тестовые данные для параметризованных тестов решено было хранить в tsv-файлах, статичные тестовые данные — в yaml. Принадлежность тестовых данных к тесту и локали решили определять при помощи названий и иерархии директорий, в которых эти данные находятся. Наследование ведется от основной базовой локали «en-us», путём удаления, добавления уникальных данных. Также в тестовых данных возможно использовать ключевое слово «skip» и «comment» — для отмены запуска конкретного тестового случая с указанием причины. Такое наследование, например, если необходимо использовать одни и те же данные для всех локализаций сайта, то наследование происходит автоматически без каких-либо дополнительных параметров. Кстати, для данных конфигураций теста (окружение, время ожидания и т.д.) также реализовали наследование, но уже не на основе локализации, а на основе наследования глобального конфигурационного файла, конфигами тестов.
Ещё один нюанс
Получая первые отчеты, начали думать, как удобнее отображать результаты тестов в разрезе локалей. Мы посчитали, что наиболее удобно для нас разделять результаты по принципу — каждой локали свой экземпляр allure-отчета. А для агрегации общей информации по локалям мы быстренько написали простенькую, но симпатичную обёртку.
Последнее, что омрачало нашу радость, это случаи зависания отдельных запусков в тестовой среде. Забыл упомянуть, что в качестве среды для тестирования мы решили использовать классический Selenium (как окружение, наиболее приближенное к реальному). При большом количестве проверок в selenium сбоев не избежать. А также всеми «любимые» ложные срабатывания, они очень затрудняют непрерывную интеграцию и «зеленый сигнал светофора» для продакшн.
Подумали и нашли выход. Зависания — побороли доработкой нашей обертки. Добавили возможность указать максимальное время выполнения отдельного теста и, если он не выполняется за указанное время, мы его грубо перезапускаем. А ложные срабатывания убрали при помощи дополнения rerun-xfails для pytest. Этот плагин автоматически перезапускает все проваленные тесты, количество попыток опять же задаем в конфигурационном yaml-файле для каждого теста или общий.
Эпилог
И, наконец, вот оно — счастье начинающего автоматизатора: стабильная удобная рабочая система. Она проста в обслуживании, позволяет провести тестирование максимально быстро и без ложных срабатываний, предоставляет очень удобную отчетность о результатах тестирования.
P.S.
Друзья, по вашим фидбекам здесь на Хабре мы бы хотели понять, насколько интересен наш опыт. Есть идея опубликовать получившееся готовое решение в виде docker-контейнера.
© Habrahabr.ru
Дефекты продукта и производительность
Многие люди по эту сторону океана с зевотой встретили шумиху по поводу чудесных достижений Японии в производстве за последнее десятилетие. «Так что же тут нового?» они говорят. «Деминг сказал это много лет назад». У. Эдвардс Деминг — если не настоящий отец, то крестный отец продуктивности растений. Он в основном отвечал за внедрение на заводах процедур статистического контроля, которые так ловко переняли японцы.В этой статье описывается подход Деминга к производительности и качеству: поскольку только руководство имеет право изменять производственную систему для их устранения, дефекты продукции являются ответственностью руководства.
Чтобы изменить систему, руководство сначала должно отличить ненормальное от нормального отклонения. Также необходимо оперативно указать, что должна производить система. Имея эти средства контроля, организация может прогнозировать производительность, затраты и уровни качества, а менеджеры могут эффективно общаться с клиентами и людьми в цехе.И это наиболее важно, поскольку, когда руководство видит причину проблем в системе, а не в рабочих, исчезают многие из подрывающих моральный дух результатов плохих решений, целей, в которые никто не верит, и мотивирующих лозунгов, косвенно обвиняющих рабочих.
Джон Генри, президент Global Manufacturing Company, откинулся на спинку стула, вздохнул и уставился в потолок. На столе перед ним лежал отчет двух статистиков о проблемах производительности и качества на фабрике Global’s Nightingale.
Генри и его вице-президенты знали, что дела идут плохо. Клиенты жаловались, цены были слишком высокими, дебиторская задолженность росла, звонки на ремонт росли, затраты росли, моральный дух рабочих падал, а профсоюз грозил забастовкой из-за постоянных требований руководства к повышению производительности. Кроме того, большинство машин было не для работы. Но они не договорились, с сожалением подумал Генри, о том, что найдут статистики. Он взял отчет, вздохнул в другой раз и снова посмотрел на него.
«Ваша фабрика в Nightingale, — говорится в отчете, — работает изо дня в день, отправляя товары, 15% из которых (в среднем) имеют один или несколько серьезных дефектов … Эта доля серьезных дефектов в вашем продукте вполне может объяснить некоторые из ваших проблем с продажами и прибылью. Количество переделок, которые операторы должны выполнять на производственной линии, также снижает вашу прибыль.
«Так начинаются ваши проблемы. Оператор на линии превращает элемент. Она смотрит на это. Если она обнаружит серьезный дефект, она может исправить его сама, потому что знает, что в противном случае он может вернуться к ней позже, чтобы исправить.Но, думает она, инспектор, возможно, не заметит этот дефект. Если она это сделает, она может переделать его или отправить обратно оператору. Но даже если инспектор увидит это, супервайзер может перехватить предмет на обратном пути к оператору и отправить его на производство, чтобы избежать короткого замыкания на более позднем этапе производства.
«Почему бы с точки зрения оператора не рискнуть и с мелкими, и с серьезными дефектами? Отправьте их по очереди; скорее всего, они не вернутся.С точки зрения инспектора, руководитель может вмешаться, поэтому исправление дефектов может быть пустой тратой времени. С точки зрения руководителя, она может рискнуть дефектом. Она не может проиграть и может выиграть, если будет поддерживать высокие производственные рекорды.
«Другими словами, мистер Генри, работа вашего оператора — устранять дефекты. Ей платят за них. Это система, и оператор не несет за нее ответственности. Управление есть ».
Суть записки статистиков бедному Джону Генри заключается в том, что дефектов не бесплатны: кто-то делает дефекты и получает деньги за их изготовление.Если значительная часть рабочей силы исправляет дефекты, то компания платит как за исправление дефектов, так и за их устранение. Если фабрика Nightingale производит 15% бракованной продукции, то 15% от общей стоимости тратится на изготовление плохих единиц. Очевидно, что низкое качество означает высокую стоимость.
Все проблемы Генри и его вице-президентов возникли из-за неправильного управления качеством. Другими словами, и это могло бы быть вторым моментом статистиков, менеджмент достигает высококачественного продукта за счет улучшения процесса.Если менеджеры смогут улучшить производственный процесс, они смогут перенаправить ресурсы с производства дефектных продуктов на производство дополнительных хороших продуктов.
Предположим, что руководство компании Nightingale способно улучшить процесс, внеся некоторые изменения без дополнительных затрат, так что только 9% выходных данных являются дефектными. Чего удалось достичь менеджменту?
1. Производительность повысилась. Теперь фабрика производит на 6% больше единиц по той же цене. (Если завод исправляет дефекты, то операторы могут использовать время, которое они потратили бы на переработку 6% дефектов, чтобы производить более качественный продукт.Это дает дополнительное — бесплатное — повышение производительности.)
2. Совокупное качество улучшилось. Теперь неисправно только 9% продукции вместо 15%.
3. Емкость увеличилась. Завод производит на 6% больше хороших единиц с той же системой — рабочая сила, машины, материалы и так далее.
4. Стоимость единицы ниже. Фабрика производит больше единиц по той же цене.
5. Цену можно снизить.
Видно, что управление процессом (т.д., надлежащее управление качеством) может облегчить проблемы Джона Генри. С повышением качества клиенты перестанут жаловаться, и возврат упадет, продавцы смогут эффективно конкурировать за счет более качественного продукта и более низкой цены, количество обращений за обслуживанием и ремонтом сократится, дебиторская задолженность снизится (потому что, скорее всего, довольные клиенты будут платить по счетам), расходы снизятся, производительность возрастет, профсоюз перестанет угрожать забастовкой, а у руководства появится капитал для надлежащего обслуживания оборудования.
Улучшение процесса — ключ к повышению производительности и качества, а также к снижению затрат на единицу продукции. Менеджеры могут достичь этих целей, понимая источники вариаций в процессе и используя соответствующие операционные определения.
Источники вариаций
Давайте посмотрим на производственный процесс, при котором производятся стальные стержни. Хотя средний диаметр стержней составляет 2,00 дюйма, мы не можем ожидать, что диаметр каждого стержня будет точно таким. Мы ожидаем некоторых вариаций в зависимости от того, как округлить результат измерения.
Изменения в процессе естественны. Фактически, мы все должны этого ожидать и не удивляться, когда это произойдет. Но процессы подвержены двум источникам вариаций: нормальным и ненормальным. Ненормальные отклонения возникают по особой или конкретной причине и могут присутствовать или не присутствовать в процессе. В нашем примере предположим, что мы производим стержень диаметром 1,96 дюйма. Несоответствие 0,04 дюйма является ненормальным изменением процесса? Или это нормальный вариант, которого мы должны ожидать? Если это ненормальное изменение, мы хотели бы вмешаться и, скажем, отрегулировать машину.Если это не так, нам не следует вмешиваться. Фактически, регулируя машину без причины, мы рискуем вывести процесс из строя.
По оценкам некоторых исследователей, аномальные отклонения вызывают 15% проблем в процессе, а нормальные отклонения вызывают оставшиеся 85%. 1 Нормальные вариации присущи всем элементам процесса — целой группе рабочих, целому отделу и даже целой компании — и создают большую часть высоких затрат на производство и обслуживание и проблемы с низкой производительностью.Путаница между общими и частными причинами вариаций приводит к разочарованию на всех уровнях, большему разнообразию и более высоким затратам. Не имея возможности различить два источника отклонений, руководство может отреагировать, обвиняя рабочих.
Рабочий бессилен устранить обычную причину изменения. У рабочих нет полномочий уточнять определения и тесты, определяющие приемлемое качество. Они мало что могут сделать с вышедшими из строя машинами или испытательным оборудованием. Они могут сообщать о таких событиях, но руководство должно следить за ними и вносить необходимые изменения.Рабочие также не могут изменять спецификации и политику закупки входящих материалов, и они не несут ответственности за дизайн продукта. Все они являются частью системы, и только менеджеры могут изменять систему.
Трудно переоценить, насколько высок моральный дух на большинстве заводов, если бы руководство возлагало на рабочих ответственность только за то, что они могли контролировать, а не за недостатки системы.
Что это за вариация?
Поскольку работники не могут нести ответственность за систему, менеджеры должны уметь различать ненормальные и нормальные отклонения, чтобы они знали, когда и как изменить процесс.Единственный безопасный способ различить два источника вариаций в процессе — использовать статистические сигналы, генерируемые контрольными диаграммами.
Контрольные карты
Контрольная диаграмма системы имеет центральную линию, которая представляет собой среднее значение процесса, и два контрольных предела, верхний и нижний. Предположим, вы хотите изучить операцию набора клавиш в отделе обработки данных. Во-первых, согласно статистической теории, вы определяете размер выборки, скажем, 200 карточек в день. 2 Затем вы берете случайные выборки из 200 карточек из ежедневных выходных документов и проверяете их на наличие ошибок.На рисунке I показано, как построить контрольную диаграмму для операции нажатия клавиш.
Приложение I Формулировка контрольной диаграммы для работы с клавиатурой Примечание. Обе точки (день 8 и день 22), которые находятся над UCL, посылают статистический сигнал руководству для поиска возможных источников аномальных колебаний в день 8 и день 22.
Приложение Ia показывает процент дефектных перфокарт. Приложение Ib представляет собой график зависимости «процента дефектных» (столбец 4 в Ia) от «дня» (столбец 1 в Ia). Приложение Ic показывает вычисления, которые потребуются вам для построения центральной линии (в этом примере — средний процент дефектов процесса), а также верхнего и нижнего контрольных пределов.
Вы создаете контрольную диаграмму (идентификатор приложения) , соединив точки, нанесенные на график Ib, и проведя через эти точки центральную линию, а также верхний и нижний контрольные пределы. Наконец, вы анализируете контрольную диаграмму. Если значение выборки попадает в верхний и нижний контрольные пределы и если тренд или какой-либо другой систематический образец отсутствует, вариация, вероятно, нормальная.Если, однако, значение выборки выходит за контрольные пределы, вероятно, отклонение ненормально.
Диаграмма, показанная на Приложении I, является лишь одним из многих видов контрольных диаграмм, каждая из которых имеет особое назначение. (Вы можете найти примеры других графиков в источниках, перечисленных в конце статьи.)
Если отклонение ненормальное
Сравнивая Ib и Id, читатель увидит, насколько сложно невооруженным глазом различить две причины вариации.Приложение Ib не позволяет менеджерам различать два источника отклонений, в то время как Exhibit Id ясно показывает, что в дни 8 и 22 произошло что-то ненормальное, не связанное с системой, что привело к перфорации дефектных карт.
Когда менеджер определяет, что причина отклонения является ненормальной, он должен искать и устранять причины, относящиеся к конкретному рабочему или группе рабочих, машине, новой партии сырья и так далее. Как только руководство устраняет все приписываемые причины отклонений, остается стабильный процесс, находящийся под статистическим контролем.
Давайте еще раз рассмотрим операцию перфорирования клавиш, показанную на Приложении I. Посмотрите на контрольную диаграмму процент карточек с ошибками (Id).
Принято основывать контрольные пределы на кратной стандартной ошибке. Обычно это кратное 3, и пределы называются пределами 3-сигма. Это означает, что есть приблизительно 3 шанса из 1000, что расположение точки вне пределов обусловлено естественным случайным изменением системы. Если мы посмотрим на графики в Приложениях I и II, мы увидим, что две точки находятся за пределами верхнего контрольного предела, что указывает на то, что процесс не находится в статистическом контроле.
Приложение II Контрольные диаграммы для клавишных перфораторов
Каким должен быть следующий шаг менеджменту? Чтобы взять процесс под контроль, руководство должно исследовать моменты, которые вышли из-под контроля, чтобы удалить из процесса назначаемые причины отклонений. Предположим, что руководство обнаружило, что на 8-й день к персоналу был добавлен новый оператор по нажатию клавиш, и что один день, который потребовался работнику, чтобы приспособиться к новой среде, вероятно, вызвал необычно большое количество ошибок с клавиатурой.Чтобы гарантировать, что эта назначаемая причина не повторится, компания ввела однодневную программу обучения.
Расследование дня 22 показало, что накануне вечером в отделе закончились карточки от обычного поставщика, и он не ожидал новой поставки до утра 23 дня. Следовательно, отдел закупил однодневный запас карточек у нового поставщика. . Руководство обнаружило, что эти карты были низкого качества, что приводило к большому количеству ошибок при перфорации клавиш. Чтобы исправить это назначаемое отклонение, руководство ввело пересмотренную политику инвентаризации и оперативно определило приемлемое качество перфокарт.
После исключения дней, для которых были обнаружены приписываемые причины отклонений, менеджеры пересчитали статистику контрольной диаграммы:
Приложение IIb показывает исправленную контрольную диаграмму (IIa показывает исходную диаграмму). Теперь процесс стабилен, находится под статистическим контролем.
Стабильный процесс, который демонстрирует только вариации из-за внутренних ограничений системы, позволяет менеджеру определять его возможности, то есть то, что является нормальным. Вот некоторые из преимуществ достижения стабильного процесса:
1.Руководство знает возможности процесса и может спрогнозировать его производительность, затраты и уровни качества.
2. При существующей системе производительность максимальна, а затраты минимальны.
3. Руководство может измерять влияние изменений в системе с большей скоростью и надежностью.
4. Если руководство хочет изменить пределы спецификации, у него есть данные для резервного копирования своего аргумента.
Возможности процесса становятся данностью. Стабильный процесс, который приводит к недопустимому количеству дефектов, будет продолжать работать до тех пор, пока система, как она определена в настоящее время, остается неизменной.И только руководство несет ответственность за изменение системы.
Нормальное изменение
Когда процесс достигает стабильности, что является не естественным состоянием, а достижением, руководство готово воздействовать на систему для повышения производительности и качества. Менеджеры могут улучшить систему:
1. Сдвиг среднего значения процесса. Например, руководство может захотеть уменьшить процент дефектов или увеличить средний результат.
2. Изменение степени вариации.Учитывая экономические требования рынка, руководство может захотеть уменьшить количество вариаций, чтобы получить более стабильно однородный продукт, или увеличить его, чтобы получить менее однородный продукт.
Определенные входные данные и процедуры, такие как труд, обучение, надзор, сырье, машины и рабочие определения, определяют систему. Чтобы улучшить систему, руководство должно изменить эти факторы. Мы снова подчеркиваем, что только руководство несет ответственность и имеет право вносить эти изменения.Сами по себе рабочие не могут повлиять на систему.
Как руководство может изменить процесс ввода клавиш для повышения производительности и качества? Внедряя процедуры обучения, которые снижают средний процент дефектных карточек и количество общих отклонений (что приводит к более узким контрольным пределам), руководство может помочь сотрудникам последовательно выпускать больше безошибочных карточек.
Приложение IIc показывает новую контрольную диаграмму после того, как руководство ввело обучение и процедурные изменения.Средний процент перфокарт с ошибками снизился с 0,017 до 0,008, а также уменьшились вариации процесса.
Важно подчеркнуть, что обсуждаемые нами концепции охватывают не только контрольные диаграммы. Компании могут использовать контрольные карты без какого-либо понимания подхода, который нас интересует, а именно ответственности руководства за улучшение системы, отсутствия привычной зависимости от окончательной проверки, исключения лозунгов, устранения произвольных стандартов работы и так далее.
Круг замкнулся. Мы знаем, что улучшение процесса увеличивает производительность и качество. Различая аномальные и нормальные отклонения и устраняя аномальные отклонения, менеджеры могут получить статистический контроль. Но одного этого недостаточно для повышения производительности и качества.
Если бы руководство полностью понимало источники отклонений, а также осознавало свою ответственность за улучшение процесса, но не понимало операционных определений, его усилия все равно были бы напрасными.
Что производится?
Если руководство не может точно определить свои продукты, как оно может их продать, описать, чего оно хочет, людям в цехе или улучшить производственный процесс? Не может. Без операционного определения люди не могут вести бизнес. Вот пример путаницы, которую может вызвать отсутствие точного представления о том, что производится:
«Этикетка на одеяле гласит« 50% шерсть ». Что это означает? Полушерсть в среднем поверх этого одеяла или полушерсть за месяц производства? Что такое полушерсть? Половина по весу? Если да, то при какой влажности? Каким методом химического анализа? Сколько анализов? Нижняя половина одеяла шерстяная, а верхняя — что-то другое? Это 50% шерсть? Означает ли 50 процентов шерсти, что в любом случайном сечении должно быть немного шерсти размером в полдоллара? Если да, сколько разрезов следует проверить? Как вы их выбираете? Какому критерию должно удовлетворять среднее значение? И насколько допустимы отклонения между разрезами? Очевидно, что значение 50% шерсти можно определить только в статистических терминах.” 3
Каково точное или истинное определение термина? Например, что такое «точно круглый»? Не существует единого определения, которое помогло бы нам определить, действительно ли что-то круглое. Словарь тоже не поможет. Вебстер говорит, что фигура является круглой, если у нее «каждая часть поверхности или окружности равноудалена от центра». Это определение очень полезно для формальной логики, но если мы попытаемся использовать его, чтобы определить, круглый ли наш диск, мы столкнемся с непреодолимой трудностью. Словарь дает понятие, а не определение для использования в промышленности.
Как же тогда мы можем определить термин, понятный на уровне магазина? Операционные определения бывают двух типов: одно для атрибутов, например, успех или неудача, и другое для переменных, например, объема продаж. Рабочее определение атрибута состоит из:
1. Критерий, применяемый к объекту или группе.
2. Процедура выбора исследуемого объекта.
3. Операция, например измерение или наблюдение за объектом.
4.Запись результата.
5. Тест объекта на соответствие критерию.
6. Решение «да» или «нет» относительно того, соответствует ли объект критерию.
Чтобы получить операционное определение переменной, менеджеры предприняли бы те же первые четыре шага, которые они предприняли для получения определения атрибута. (Шаги 5 и 6 для атрибутов не применяются к переменным.)
Теперь возникает вопрос: какое значение имеют операционные определения для производительности компании? Мы знаем, насколько важно, чтобы производители и пользователи понимали друг друга.Без рабочих определений спецификация бессмысленна. Конфликты и неразбериха между компаниями и между отделами компании возникают из-за того, что менеджеры не могут заранее в значимых терминах указать спецификации элемента или его характеристики. Подумайте о проблемах производительности и качества, которые могут возникнуть, когда инспектор, ответственный за обнаружение дефектов, со временем не согласуется в своих суждениях или когда инспекторы противоречат друг другу. Рабочие не знают, что приемлемо, а что дефектно.Им нужно оперативное определение бракованного продукта.
Допустим, мы производим круглые диски. Диски круглые? Почему нас это волнует? Если диск слишком далек от круглой формы, это приведет к заклиниванию машины клиента, повреждению оборудования и простоям. Если мы хотим оставаться в бизнесе, о нас лучше заботиться.
Запишем рабочее определение раунда для диска. Поскольку мы измеряем атрибут (круглое или не круглое), мы будем работать над первым типом операционного определения.
Шаг 1: Сначала мы хотим вывести критерий для объекта.
а. «Используйте штангенциркуль в достаточно хорошем состоянии». (Вы сразу чувствуете необходимость подвергать сомнению каждое слово.)
«Что такое« достаточно хороший порядок »?» (Мы решаем вопрос, позволяя вам пользоваться штангенциркулем.)
«Но как мне их использовать?»
«Мы будем довольны, если вы будете использовать их как обычно».
«При какой температуре?»
«Температура в этой комнате.”
г. «Возьмите 6 мерок диаметром примерно 30 градусов друг от друга. Запишите результаты ».
«Но что такое« примерно 30 градусов друг от друга »? Разве вы не имеете в виду ровно 30 градусов? »
«Нет, в физическом мире не бывает ровно 30 градусов. Так что попробуйте на 30 градусов; мы будем довольны ».
г. Если диапазон между 6 диаметрами не превышает 0,007 сантиметра, мы объявляем диск круглым. Мы определились с критерием.
Шаг 2: Давайте выберем конкретный диск.(На этом этапе мы могли бы указать некоторую схему выборки.)
Шаги 3 и 4: Произведите измерения и запишите результаты в сантиметрах — 3,365, 3,363, 3,368, 3,366, 3,366 и 3,369.
Шаг 5: Диапазон от 3,369 до 3,363, или 0,006 разницы. Мы проверяем соответствие, сравнивая диапазон 0,006 с диапазоном критерия меньше или равным 0,007 (из шага 1).
Шаг 6: Поскольку диск прошел предписанный тест на округлость, мы объявляем его круглым.
Если в компании есть работники, которые понимают, что означает раунд, и заказчик, который соглашается, проблемы, которые могли возникнуть у компании, чтобы удовлетворить клиента, исчезнут.
Давайте посмотрим на другой пример, где операционные определения улучшают понимание внутри компании. В этом примере мы измеряем переменную (продажи), поэтому мы используем второй тип операционного определения.
Продавцу говорят, что о ее работе будут судить по проценту изменения продаж в этом году по сравнению с продажами в прошлом году.Что это значит? Среднее процентное изменение каждый месяц? Каждую неделю? Каждый день? Для каждого продукта? Процентное изменение продаж с 31 декабря 1980 г. по 31 декабря 1981 г.?
Как мы измеряем продажи: валовую, чистую, валовую прибыль, чистую прибыль и т. Д.? Процентное изменение выражено в постоянных или завышенных долларах? Если это в постоянных долларах, каков базовый год? Если оно в завышенных долларах, то по ценам прошлого года или этого года? В каких экономических условиях?
Расплывчатое определение процентного изменения может привести только к путанице, разочарованию и недоброжелательности между менеджментом и торговым персоналом — вряд ли способ повысить производительность.Как руководство должно оперативно определять процентное изменение продаж?
Шаг 1: Процентное изменение продаж — это разница между продажами 1981 г. (с 1 января 1981 г. по 31 декабря 1981 г.) и продажами 1980 г. (с 1 января 1980 г. по 31 декабря 1980 г.), деленная на объем продаж 1980 г .:
S80 измеряется в постоянных долларах, с 1979 годом в качестве базового, с использованием цен на 15 июня 1979 и 15 июня 1980 года для получения постоянных долларовых цен и общего объема продаж единицы продукции за вычетом прибыли (по любой причине) по состоянию на 31 декабря. 1980 г. для каждого продукта.
S81 измеряется в постоянных долларах с 1979 г. в качестве базового года с использованием цен на 15 июня 1979 г. и 15 июня 1981 г. для получения постоянных долларовых цен и общего объема продаж единицы продукции за вычетом прибыли (по любой причине) по состоянию на 31 декабря 1981 г. для каждого продукта. (Pi79 остается неизменным для всех продуктов.)
Эта процедура для расчета процентного изменения продаж в период с 1980 по 1981 год будет действовать независимо от экономических условий. Кроме того, руководство может пересмотреть определение процентного изменения продаж после оценки продаж 1985 года, но не раньше, если торговый персонал и руководство продаж не согласятся.
Шаг 2: Продавец и ее записи о продажах являются объектом исследования.
Шаги 3 и 4: Менеджер по продажам будет использовать все счета-фактуры за 1980 и 1981 гг. И квитанции о продажах для вычисления чистого количества единиц, проданных для каждого продукта в 1980 и 1981 гг.
Предыдущее определение продаж может не подходить другому менеджеру или продавцу; однако, если менеджер по продажам принимает его, а сотрудники отдела продаж понимают его, это операционное определение.
Операционные определения нетривиальны. Если руководство не определит многие критические переменные и атрибуты так, чтобы их согласовали и работники, и клиенты, последуют серьезные проблемы. Контрольная диаграмма становится бесполезным управленческим инструментом из-за совершенно нового источника вариаций: вариаций измерения. Оперативное определение характеристик, отображаемых на карте, является обязанностью руководства. Если инспекторы не соглашаются друг с другом или с самими собой изо дня в день, разовьется хаос.Рабочие не знают, чего от них ждут. Их вывод подходит для Инспектора 1, но не для Инспектора 2; работа сотрудника могла быть сдана инспектором 1 вчера, но не сегодня.
Должность высшего руководства
Ряд людей недавно написали руководящие принципы, которые говорят руководству, что ему следует делать для повышения производительности:
Создайте организацию с постоянной целью и долгосрочными обязательствами высшего руководства.
Разрушайте барьеры между отделами.
Создайте среду, в которой люди не боятся сообщать о проблемах.
Обезвредить встроенные уровни дефектов, ошибок, некачественных материалов и т. Д.
Не вините рабочих в проблемах производительности и качества.
Читатель, несомненно, знаком с ними. Однако из изложенного здесь подхода вытекают дополнительные руководящие принципы, которые могут быть не столь очевидными.
1. Не ждите, что проверка решит проблему качества. К моменту проведения проверки товар уже годен или неисправен. Вы не можете проверить качество продукта.
Массовая проверка не позволяет четко отделить хорошие предметы от плохих. Лучшим способом является наблюдение за небольшими образцами продукта для контрольных диаграмм для достижения или поддержания статистического контроля. Таким образом менеджеры могут избавиться от необходимости инспектировать и использовать таланты инспекторов для других целей. Продавцы и покупатели также могли сравнить свои инструменты и тесты; продавцы и покупатели могли начать говорить на одном языке.Осмотр под давлением часто оказывается фарсом: идет он или уходит, все проходит. А поскольку разделение ответственности означает, что никто не несет ответственности, 200% проверка менее надежна, чем 100% проверка.
2. В соответствии с политикой, прекратите предлагать бизнес лицу, предложившему самую низкую цену. Цена без меры качества не имеет значения.
Чтобы судить о качестве, менеджерам по закупкам требуется образование и опыт в оценке статистических свидетельств качества.Если покупатели станут экспертами в оценке качества, большинство из них резко сократит количество поставщиков, с которыми они имеют дело. Поставщик, который не знает ни своих затрат, ни того, сможет ли он повторить сегодняшнее распределение качества завтра, не является хорошим деловым партнером.
3. Избавьтесь от целей, числовых целей, лозунгов («ноль дефектов»), изображений и плакатов, которые руководители так часто наклеивают на предприятиях, призывая людей повышать производительность. К сожалению, такие программы «повышения производительности» оставляют дефекты там, где они есть.Они не выявляют и не исправляют неисправности системы, а также не предоставляют менеджерам статистических сигналов, которые необходимы для принятия корректирующих действий. Они не отвечают на критический вопрос: «Как мы можем повысить производительность?»
4. Отменить квоты на работу. Рабочие квоты не учитывают обычные изменения в системе. Они не включают способ обнаружения необходимости в корректирующих действиях или способ возложить ответственность на руководство или на представителя руководства на линии. Например, менеджер банка может определить количество клиентов, которые, по его мнению, кассир должен обработать за час, количество вычислений процентов и штрафов, которые кто-то должен вычислить за час, и аналогичный показатель для всех остальных действий.Однако стандарты ничего не говорят о качестве работы и не дают руководителю возможности понять различия в процессе. Стандарты не указывают, какие действия следует предпринять менеджерам или как улучшить процесс.
5. Разработайте учебные программы по статистике, чтобы менеджеры и руководители могли понять, как управлять качеством. Надзор является частью системы и, разумеется, является обязанностью руководства. Статистические методы являются жизненно важными помощниками для мастеров и руководителей производства в выявлении причин потерь, низкой производительности и низкого качества.Менеджеры также могут использовать их, чтобы определить, когда сотрудники полностью обучены и когда поможет дальнейшее обучение.
• • •
Настоящее руководство указывает, что высшее руководство должно делать для повышения производительности и качества. Хотя следование каждому из руководящих принципов не принесет ощутимых результатов, в то же время компания, начавшая сегодня с полной отдачей, вскоре добьется впечатляющих результатов.
Ближайшая секунда для быстрых результатов — это избавление от страха, помощь людям в безопасности и преодоление страха сообщать о проблемах с оборудованием или поступающими материалами.Менеджеры могут достичь этой цели за два-три года и получить серьезные экономические результаты.
1. См., Например, Джозеф М. Джуран, Справочник по контролю качества , 3-е изд. (Нью-Йорк: McGraw-Hill, 1974).
2. Обсуждение того, как вычислить размер выборки, можно найти в многочисленных текстах, некоторые из которых мы перечислили в конце статьи.
3. У. Эдвардс Деминг, Качество, производительность и экономическое положение (Кембридж: Центр перспективных разработок Массачусетского технологического института, 1982 г.).
Версия этой статьи появилась в выпуске Harvard Business Review за сентябрь 1983 г.Внедрение полностью интегрированной системы управления без дефектов
Сатвант Рахра. Эта статья опубликована в выпуске Inspectioneering Journal за март / апрель 2019 года.| Эта статья является второй частью серии из трех статей. |
| Часть 1 | Часть 2 | Часть 3 |
Примечание редактора: Часто говорят, что унция профилактики стоит фунта лечения. Следующая серия статей посвящена методам устранения потенциальных дефектов в оборудовании путем выявления условий, ведущих к возникновению дефектов, и их упреждающего устранения. Следовательно, инспектору будут знакомы такие методы, как IOW, инспекция и контроль качества.Это вторая часть серии, состоящей из нескольких частей. Часть 1 появилась в выпуске Inspectioneering Journal за январь / февраль 2019 года.
Введение
Ошибочное управленческое мышление в отношении систем, процессов и устойчивой практики продолжает приводить нас к созданию производственных предприятий, отказы которых начинаются вскоре после ввода в эксплуатацию. Стремление к бездефектной работе требует усилий, чтобы систематически и эффективно снижать текущую частоту отказов оборудования.Предлагаемая пятикомпонентная бизнес-стратегия, обсуждаемая в этой серии статей, по мнению автора, является противоядием от этих проблем. Стратегия включает:
- Команда менеджеров по новому мышлению
- Развивать образованную рабочую силу
- Используйте комплексную систему управления (MS)
- Внедрить лучшие процессы
- Прививайте культуру постоянного совершенствования
Стратегия представляет собой уникальное сочетание важнейших концепций и систем, реализация которых поможет руководству значительно улучшить свои операционные показатели.До этого момента эти концепции и системы обсуждались только изолированно, и поэтому любой организации было сложно установить связи и эффективно применять их в своей деятельности. Применение этого бережливого мышления и методологий нулевого контроля качества (QC) Shingo, которые активно помогают обнаруживать и устранять условия, которые могут привести к ошибкам и дефектам, являются отличительными чертами этой стратегии.
В данном документе описаны методологии нулевого контроля качества, а их применение поясняется на промышленных примерах.Таблица используется, чтобы помочь персоналу проанализировать условия, которые могут привести к созданию ошибок, возникновению ошибок и последующим последствиям. Разработан план смягчения каждой ошибки и определены сроки, чтобы гарантировать, что ни одна ошибка не перерастет в дефект. Ключевым элементом стратегии является внедрение структурированной, всеобъемлющей и интегрированной основной системы MS. Подробно рассматривается разработка основной СУ и интеграция различных мероприятий, требований и руководств, чтобы позволить любой управленческой команде разработать и внедрить СУ, подходящую для ее организации.Комплексная и полностью интегрированная основная система MS, способная предоставлять систематические и подробные инструкции, полностью поддерживаемая эффективными рабочими процессами, увеличивает вероятность достижения бездефектных процессов проектирования, закупок, изготовления, строительно-монтажных-пусконаладочных работ, эксплуатации и технического обслуживания. Рекомендации и обзор отраслевых пробелов, которые были обнаружены автором и другими полевыми сотрудниками за период двух десятилетий инспекций и аудита, представлены на протяжении всего процесса применения МС к производственным процессам.
Аспекты операции, такие как сам процесс, безопасность персонала, качество продукции, целостность оборудования и защита окружающей среды, взаимосвязаны и неотделимы друг от друга. Системы и процессы, реализованные на протяжении всей операции, вплетены в нить всеобъемлющей MS, которая лежит в основе любой операции. Достижение безупречной работы — исключительный результат, который может быть достигнут только благодаря чрезвычайным усилиям (т.е., реализация 5-компонентной бизнес-стратегии, сформулированной в этой статье). Эта статья была написана автором после выхода на пенсию и основана на его жизненном опыте работы в промышленности, изучении работ многих авторитетных авторов в области качества, целостности оборудования, работающего под давлением, бережливого мышления, практик ISO и RAGAGEP (признанных и общепринятые передовые инженерные практики).
Автор предполагает, что бездефектная работа предприятия может быть реализована, если самый старший руководитель и его / ее команда высшего руководства будут реализовывать эту бизнес-стратегию из пяти направлений :
- Развивать трансформационное понимание и понимание «бережливого мышления» среди менеджеров и создавать среду, в которой «бережливое мышление» внедряется во всей организации;
- Преобразовать всю организацию, проводя обучение по повышению осведомленности, системное обучение и обучение навыкам;
- Внедрить полностью интегрированную и задокументированную систему управления, с помощью которой можно успешно реализовать стратегию «бережливого мышления»;
- Постоянно улучшать и обеспечивать соответствие рабочих процессов заданным целям в области качества, безопасности, производства и защиты окружающей среды; и,
- Внедряйте автономную культуру совершенствования во всей организации, чтобы обеспечить постоянное улучшение возможностей, безопасности, качества, производительности и для поддержания достигнутых успехов.
Оборудование выходит из строя по причинам, неизвестным отрасли, или из-за неправильного управленческого мышления, процессов, систем управления, проектирования, закупок, строительства, монтажа, эксплуатации и / или методов технического обслуживания. Все сбои, связанные с неправильной практикой, можно предотвратить, они будут исследованы и предложены решения в этой серии статей. В первой части этой серии из трех частей автор сосредоточился на описании «бережливого мышления» и того, что это потенциально может означать для вашей работы.В Части 2 автор изложит стратегии, которые операторы могут реализовать для уменьшения количества отказов, устранения расточительной деятельности, снижения затрат, увеличения пропускной способности, повышения безопасности и достижения цели бездефектной работы.
Понимание рационального мышления
Итак, напомним, в контексте технологического предприятия состояние бездефектной работы считается достигнутым, когда организация успешно реализует 5-компонентную бизнес-стратегию, изложенную выше, предотвращает повреждение или отказ своего оборудования, выполняет действия, которые неизменно соответствует его критериям качества и производит продукт, соответствующий его спецификациям.
Этот контент доступен зарегистрированным пользователям и подписчикам.
Зарегистрируйтесь сегодня, чтобы получить доступ к этой статье бесплатно.
Создайте бесплатную учетную запись и получите доступ к:
|
Текущие подписчики и зарегистрированные пользователи могут войти в систему сейчас.
растений, вышедших из строя при переходе между состояниями, могут в значительной степени компенсировать свой недостаток | Физиология растений и клетки
Аннотация
Arabidopsis thaliana растений, лишенных субъединицы PSI-H или PSI-L фотосистемы I, серьезно страдают от их способности выполнять переходы между состояниями, но визуального фенотипа не наблюдалось, когда эти растения выращивались при разных количествах света и качества. Однако хлоропласты у растений без PSI-H и PSI-L содержали меньше и более протяженные стеки гран.Растения, лишенные PSI-H или PSI-L, были охарактеризованы в отношении их фотосинтетических способностей. Растения дикого типа отрегулировали нефотохимическое тушение флуоресценции для поддержания постоянных уровней квантового выхода ФСII и уменьшения пула пластохинонов. Напротив, растения с дефицитом переходов между состояниями имели более редуцированный пул пластохинона и, следовательно, менее эффективную фотохимию ФСII в условиях роста и освещения и в состоянии 2. Максимальная фотосинтетическая способность и квантовая эффективность выделения кислорода были уменьшены в 8–8 раз. 14% в установках без PSI-H.В условиях роста и освещения строма была аналогичным образом восстановлена у растений без PSI-H, и скорость циклического транспорта электронов не изменилась. Пигментный анализ показал, что цикл ксантофилла не активируется, чтобы компенсировать отсутствие переходов между состояниями. В целом, растения, лишенные PSI-H и PSI-L, показали пониженную способность оптимизировать фотосинтез в соответствии с условиями освещения.
(получено 9 июля 2002 г.; принято 31 октября 2002 г.)
Введение
Растения собирают свет путем кислородного фотосинтеза через фотосистему II (ФСII) и фотосистему I (ФСI), тем самым потребляя углекислый газ и высвобождая кислород.Линейный транспорт электронов через PSII и PSI катализирует перенос электронов от воды к NADP + и приводит как к образованию сильных восстановителей, так и к образованию протонного градиента, используемого для управления АТФ-синтазой. Помимо работы в связи с PSII, PSI может работать самостоятельно, выполняя циклический перенос электронов, способствуя протонному градиенту через тилакоидную мембрану. PSI представляет собой мультибелковый комплекс, состоящий из различных субъединиц у высших растений (от PSI-A до PSI-L, PSI-N и PSI-O), каждая из которых обнаруживается в виде одной копии на реакционный центр (обзор Chitnis 2001, Scheller et al.2001). Реакционный центр управляет переносом электронов от пластоцианина на просветной стороне тилакоидной мембраны к ферредоксину на стромальной стороне. PSI собирает свет через приблизительно 96 хлорофиллов, связанных с самим центральным комплексом, и посредством взаимодействия со светособирающим антенным комплексом I (LHCI) (Jordan et al. 2001). PSI-A, PSI-B и PSI-C связывают все кофакторы, участвующие в переносе электронов, и эти субъединицы являются высококонсервативными среди различных организмов, показывая, что основной механизм переноса электронов идентичен у цианобактерий, водорослей и высших растений.Однако в процессе эволюции многие организмы развили дополнительные функции для оптимизации своего фотосинтетического аппарата, например четыре субъединицы, уникальные для эукариот (PSI-G, -H, -N и -O) (Knoetzel et al. 2002).
Несмотря на то, что PSII и PSI работают последовательно, в тилакоидах пространственно разделены, PSI локализуется в не прижатых ламеллах стромы, а PSII находится в стеках прижатых гран. Две фотосистемы также имеют несколько разные спектры поглощения. Голо-комплекс PSI-LHCI содержит в основном хлорофилл a и более эффективно поглощает свет в дальней красной области, тогда как PSII, связанный с его светособирающим белком, содержит более высокую долю хлорофилла b и предпочтительно поглощает свет синего и синего цветов. красный регион.Таким образом, PSII становится ограничивающим при длине волны падающего света более 680 нм, когда PSI возбуждается преимущественно. Когда растения подвергаются освещению в пользу ФСII или ФСI, они могут перераспределять возбуждение, так что ограниченная светом фотосистема получает больше энергии. Это перераспределение энергии возбуждения регулируется посредством окислительно-восстановительного состояния пула пластохинонов (обзор Haldrup et al. 2001, Kruse 2001, Wollman 2001). Когда предпочтение отдается ФСII (состояние 2), пул пластохинона становится более уменьшенным, и это активирует киназу в тилакоидах (Allen et al.1981, Зито и др. 1999). По общему мнению, активированная киназа фосфорилирует LHCII и вызывает отслоение комплекса мобильной антенны от ФСII. Когда PSI в основном возбужден (состояние 1), растение пытается направить больше поглощенной световой энергии на PSII, что приводит к усилению возбуждения PSII. Однако роль переходов между состояниями все еще остается предметом дискуссий. Было предложено, чтобы переходы между состояниями выполняли несколько ролей, например: оптимизация поглощения света в ограниченных условиях или уравновешивание производства НАДФН / АТФ за счет увеличения циклического транспорта электронов (Wollman 2001).Однако не было обнаружено никакой оптимизации квантовых требований для фиксации CO 2 при чередовании света PSI и PSII (Andrews et al. 1993). Альтернативно, переходы между состояниями могут служить для защиты растения от фотоингибирования ФСII и чрезмерного восстановления межсистемной цепи. Недавно было показано, что киназа LHCII инактивируется восстановленным тиоредоксином в условиях высокой освещенности (Rintamäki et al. 2000). Таким образом, если фосфорилирование LHCII является основным детерминантом переходов между состояниями, то отсутствие киназной активности при ярком освещении не согласуется с фотозащитной ролью.Исследования мутантов с дефицитом перехода между состояниями Synechocystis pcc 6803 и Chlamydomonas reinhardtii до сих пор не смогли различить разные возможности. Мутант rpaC в Synechocystis pcc 6803 имеет нормальную скорость роста при высокой или средней интенсивности света, но растет значительно медленнее при слабом освещении (Emlyn-Jones et al. 1999). Поэтому было высказано предположение, что переход между состояниями у цианобактерий играет лишь незначительную фотозащитную роль и в первую очередь используется для оптимизации фотосинтетической способности.Два мутанта с дефицитом переходного состояния были идентифицированы у C. reinhardtii. Один мутант, stm1 , растет хуже, чем дикий тип как при низкой, так и при высокой интенсивности света, но в меньшей степени при умеренной интенсивности света, тогда как второй мутант, stt7 , кажется, растет так же хорошо, как и дикий тип. при любой интенсивности света (Kruse et al. 1999, Fleischmann et al. 1999). В C. reinhardtii , где 80% LHCII является мобильным и присоединяется к PSI во время перехода из состояния 1 в состояние 2, было показано, что циклический перенос электронов превышает линейный перенос электронов в состоянии 2 (Delosme et al.1996, Finazzi et al. 1999). Сомнительно, могут ли переходы между состояниями иметь аналогичную роль у растений, во-первых, количество мобильного LHCII меньше (≈20%), а во-вторых, у гороха в состоянии 2 не может быть обнаружено увеличения емкости циклических электронов по сравнению с состоянием 1 (Havaux 1992). . Следовательно, вероятно, что переходы между состояниями были оптимизированы для различных функций у про- и эукариот. Роль переходов между состояниями также может быть различной в одноклеточных и многоклеточных организмах.
Недавно мы охарактеризовали Arabidopsis thaliana растений, лишенных PSI-H, которые не могли выполнять переходы между состояниями (Lunde et al.2000). В отсутствие PSI-H поперечное сечение антенны PSI оставалось идентичным дикому типу в состоянии 1, независимо от освещения, тогда как поперечное сечение антенны дикого типа в состоянии 2 было значительно увеличено по сравнению с состоянием 1. Электронная микроскопия PSI растений показала, что LHCI связывается со стороной PSI-F / J PSI, противоположной PSI-H (Boekema et al. 2001), предполагая, что экспонированная PSI-H образует сайт связывания для LHCII на PSI. Хотя растения, лишенные PSI-H, не могут выполнять переходы между состояниями, не было обнаружено очевидных различий в их внешнем виде, когда они выращивались в оптимальных условиях в камере для выращивания.В этих условиях растения, по-видимому, компенсируют менее эффективный PSI, увеличивая соотношение PSI / PSII, на что указывает более высокое соотношение P700 / хлорофилл на 15% (Naver et al. 1999). Поэтому нас интересовало, как отсутствие переходов между состояниями влияет на физиологию растений в различных условиях освещения. Удаление PSI-H приводит к снижению субъединицы PSI-L на 40%, а растения, лишенные PSI-L, в свою очередь, обнаруживают вторичную потерю PSI-H (снижение на 80%). Эти наблюдения убедительно подтверждают тесную связь между этими двумя субъединицами PSI, как уже показали исследования перекрестного связывания (Jansson et al.1996). Поэтому в исследование были включены растения, лишенные PSI-L. Растения без PSI-L несколько меньше подвержены изменениям состояний, чем растения, полностью лишенные PSI-H, и 30% мобильного LHCII все еще чередуются между PSII и PSI при переходах между состояниями. Нарушение перехода между состояниями у растений без PSI-L наиболее вероятно из-за вторичной потери PSI-H (Lunde et al. 2000). У цианобактерий PSI-L необходим для образования тримеров (Chitnis and Chitnis 1993). Однако не было представлено никаких доказательств того, что PSI у растений собирается в тримерах (Boekema et al.2001), а функция PSI-L в растениях не выяснена.
Результаты
Антисмысловое подавление полипептида PSI-L
Фрагмент кДНК Arabidopsis PSI-L был клонирован в антисмысловой ориентации позади конститутивного промотора 35S и впоследствии трансформирован в Arabidopsis . Семена трансформированных растений собирали и проращивали на чашках, содержащих канамицин. Были выделены приблизительно 40 устойчивых к канамицину растений, которые позволили им самоопыться, и были собраны семена Т2.Четырнадцать линий Т2 проращивали на чашках, содержащих канамицин, и после переноса на торф семь растений каждой линии анализировали иммуноблоттингом на присутствие PSI-L. Было обнаружено, что в четырех из 14 линий большое количество особей имеет 0–2% PSI-L. Чтобы исследовать возможную вторичную потерю других белков PSI или LHCI, тилакоиды были выделены и проанализированы с помощью иммуноблоттинга. У растений без PSI-L было только ≈20% PSI-H, тогда как уровни других субъединиц близки к PSI-L, т.е.PSI-D, не были затронуты (рис. 1). Другие субъединицы PSI (PSI-C, D, E, F, G, K и N) и антенные белки (Lhca1-4) присутствовали в нормальных количествах.
Рост в разных условиях
Как сообщалось ранее, у растений без детектируемого PSI-H наблюдается значительный дефицит переходов между состояниями, а у растений с низким или неопределяемым количеством PSI-L, но все еще сохраняющим 20% PSI-H, наблюдается немного большее количество переходов между состояниями (Lunde et al. 2000). . Однако у растений Arabidopsis , лишенных PSI-L, не наблюдается или наблюдается лишь очень небольшое изменение внешнего вида при выращивании в постоянных и оптимальных условиях (данные не показаны).О таком же отсутствии очевидного фенотипа ранее сообщалось для растений, лишенных PSI-H (Naver et al. 1999). Кроме того, не было обнаружено никаких очевидных различий в отношении морфологии, пигментации или скорости роста, когда растения без PSI-H выращивали при слабом освещении (30 мкмоль м -2 с -1 ), высоком (400 мкмоль м –2 с –1 ) или при очень сильном освещении (1000 мкмоль м –2 с –1 ) или при повышенной температуре (30 ° C). Вероятно, что растения с дефицитом перехода между состояниями могут адаптироваться к различным, но постоянным условиям, изменяя соотношение PSI / PSII.Поэтому также тестировали рост в условиях переменного освещения. Чтобы вызвать повторяющиеся переходы между состояниями, набор растений без PSI-H и PSI-L подвергали 15-минутным циклам чередования света PSI и PSII, а другой набор — чередующемуся высокому (600 мкмоль м -2 с -1 ) и слабый (40 мкмоль м –2 с –1 ) свет. Ни в одном из этих условий визуальный фенотип не проявлялся. Пригодность растений без PSI-H при переменном освещении определяли путем подсчета количества семян, полученных на одно растение.При чередовании света PSI / PSII средний набор семян был очень похож (дикий тип 750 ± 100, PSI-H 770 ± 120, ± SE, n = 10), тогда как при чередовании высокого и низкого света количество семян уменьшалось на 10%. семян, продуцируемых растениями без PSI-H (дикий тип 1,210 ± 290, PSI-H 1,090 ± 270, ± SE, n = 10). Из-за большого разброса количества семян, производимых на одно растение, мы не можем сделать вывод о том, является ли это снижение значительным.
Тушение флуоресценции и фотосинтетическая способность
Хотя мы не наблюдали ростового фенотипа, можно предположить, что отсутствие переходов между состояниями в отсутствие PSI-H влияет на окислительно-восстановительные условия в тилакоидах.Фотохимическое тушение часто используется в качестве индикатора окислительно-восстановительного уровня первичного акцептора электронов Q A , хотя зависимость не является строго линейной (Havaux et al. 1991, Schreiber et al. 1994). Следовательно, давление возбуждения ФСII (оцененное как 1 – q P ) измерялось непосредственно в камере для выращивания в условиях освещения, к которым растения были адаптированы. В этих условиях 1 – q P увеличивался в 10 раз у растений, лишенных PSI-H, а фотохимическая эффективность (ϕ PSII ) была на 8% ниже по сравнению с диким типом ( P <10 –5 , таблица 1).Окислительно-восстановительный уровень Q A отражает баланс между восстановлением реакционным центром PSII посредством поглощения света и скоростью окисления посредством межцепочечного переноса электронов (Maxwell et al. 1995). Таким образом, даже в этих постоянных и оптимальных условиях роста поток электронов через две фотосистемы у растений дикого типа лучше сбалансирован.
Параметры флуоресценции (1 – q P , ϕ PSII , NPQ) также определялись при чередовании света PSII и PSI.В состоянии 1, когда LHCII связан с ФСII, не было никаких серьезных различий в отношении характеристик флуоресценции. Когда растения подвергались воздействию света состояния 2, давление возбуждения было примерно в два раза выше для растений без PSI-H и PSI-L ( P <10 –14 и 10 –8 , соответственно, Таблица 1). Т.о., более медленный транспорт электронов через PSI приводит к более восстановленному пулу пластохинона под светом PSII (Lunde et al. 2000). Кроме того, мы обнаружили только увеличение нефотохимического тушения у дикого типа при изменении света из состояния 1 в состояние 2 света ( P <3 × 10 –5 ).Квантовый выход фотохимии ФСII, ϕ ФСII, был лишь немного снижен у дикого типа при переходе из состояния 1 в состояние 2, тогда как значительное снижение наблюдалось как у растений без PSI-H, так и без PSI-L ( P <10 –12 и 10 –6 соответственно). Следовательно, хотя соотношение PSI / PSII было повышено у растений, лишенных PSI-H (Naver et al. 1999), они все же испытывали повышенное восстановление пластохинона и снижение ϕ PSII .
Кривые светового отклика для флуоресценции хлорофилла были также получены для того, чтобы определить реакцию растений без PSI-H по сравнению с диким типом в условиях высокой освещенности (рис.2). У растений дикого типа и без PSI-H не было обнаружено различий в эффективности фотохимии ϕ PSII , и не было обнаружено никаких серьезных изменений в NPQ. Однако NPQ растений без PSI-H действительно имеет тенденцию сдвигаться с более низкого уровня, чем у дикого типа, ниже 300 мкмоль м –2 с –1 , чтобы стать выше, чем у растений дикого типа при высокой интенсивности света. Растения, лишенные PSI-H, показали немного более окисленное состояние Q A выше плотности потока фотонов (PFD) 150 мкмоль м -2 с -1 , на что указывает более низкий 1 – q P ( P <0.04). Это было большим сюрпризом и отражает различный спектральный состав актиничной ксеноновой лампы и света камеры выращивания, причем последняя содержит меньше красного света.
Чтобы изучить влияние фотосинтетической способности растений, лишенных PSI-H, было определено выделение кислорода в зависимости от интенсивности света (рис. 3). Чтобы рассчитать квантовый выход на основе поглощенного света, были измерены пропускание и отражение света. И листья дикого типа, и листья PSI-H отражали 0.11 ± 0,01 (± SE, n = 15) доля света, которому они подвергались. Было определено, что пропускание света составляло 0,09 и 0,10 (SE ± 0,01, n = 15–18) для растений дикого типа и растений без PSI-H, соответственно. Таким образом, нет значительной разницы в количестве поглощенного света диким типом и растениями, лишенными PSI-H. Квантовый выход был рассчитан из линейного увеличения выделения кислорода от 0 до 60 мкмоль фотонов m –2 с –1 . Наклоны были 0.116 ± 0,004 (± SE, n = 18) и 0,107 ± 0,002 мкмоль O 2 (мкмоль фотонов) –1 (± SE, n = 18) для дикого типа и без PSI-H растения соответственно. Разница значительная ( P <0,05). Это означает, что растения без PSI-H используют слабый свет примерно на 8% менее эффективно, чем растения дикого типа. При ярком освещении (> 160 мкмоль м –2 с –1 ) растения без PSI-H также показали меньшие показатели с выделением кислорода на 14% ниже, чем растения дикого типа ( P <0.04). Аналогичный, но менее выраженный эффект наблюдался с растениями без PSI-L (снижение ≈5%). Интересно, что точка компенсации света составила всего 4,5 ± 0,6 мкмоль м –2 с –1 (± SE, n = 18) в растениях без PSI-H. Это значение намного ниже, чем 7,3 ± 0,7 мкмоль м –2 с –1 (± SE, n = 18), найденное для дикого типа ( P <0,01). Поведение светонасыщенных растений без PSI-H напоминает поведение теневых листьев.Однако поперечные срезы листьев не показали соответствующего морфологического изменения, и не было обнаружено различий в содержании хлорофилла на площади листа (данные не показаны).
Редокс-условия в строме и циклический перенос электронов
Ранее было показано, что эффективность фотовосстановления NADP + in vitro снижалась на 40–45% в отсутствие PSI-H, предположительно в результате менее эффективного взаимодействия с ферредоксином (Naver et al.1999). Поскольку эксперимент проводился в аэробных условиях, считалось возможным, что большее количество электронов ускользает от ферредоксина, например, в кислород. Таким образом, скорость фотовосстановления NADP + изолированными тилакоидами определялась как в аэробных, так и в анаэробных условиях. Для дикого типа скорость восстановления NADP + составляла 17,9 ± 0,4 и 18,4 ± 0,6 мкмоль NADPH мкмоль P700 –1 с –1 , а для растений без PSI-H 10,3 ± 0,01 и 10.4 ± 1.0 мкмоль НАДФН мкмоль P700 –1 с –1 в аэробных и анаэробных условиях соответственно. Следовательно, на восстановление НАДФ + не влияет присутствие кислорода и, следовательно, оно не связано с уходом электронов в кислород. Мы считали, что менее эффективный перенос электрона на ферредоксин может влиять на окислительно-восстановительные условия на стороне стромы. Уровень активации активности NADP + -малатдегидрогеназы измеряли для того, чтобы определить, меньше ли восстановлена строма растения без PSI-H.Изменение окислительно-восстановительных условий стромы при освещении будет отражаться на уровне активации NADP + -малатдегидрогеназы, поскольку этот фермент содержит окислительно-восстановительные дисульфидные связи и активируется через систему световой активации ферредоксин-тиоредоксин (Ashton et al., 2000). , Miginiac-Maslow et al. 2000). Кроме того, активность контролируется соотношением НАДФ + / НАДФН. Листья собирали через 15 минут, 2 часа и 4 часа от начала воздействия света и измеряли активность NADP + -малатдегидрогеназы.Образцы также подвергали воздействию сильно восстанавливающих условий и определяли общую емкость фермента. Растения с дефицитом дикого типа и переходного состояния показали аналогичное увеличение процентного содержания активированной NADP + -малатдегидрогеназы после воздействия света, и они достигли аналогичного устойчивого уровня через 2 часа (рис. 4). Это показывает, что нет разницы в окислительно-восстановительных уровнях стромы. Менее эффективный транспорт электронов к ферредоксину может также влиять на способность растений без PSI-H осуществлять циклический транспорт электронов.Таким образом, была определена кинетика восстановления P700 + после дальнего красного освещения. Уменьшение поглощения из-за уменьшения реакционного центра можно описать как сумму двух экспоненциальных распадов с одинаковой амплитудой. Для дикого типа и растений, лишенных PSI-H, k fast составляло 1,3 ± 0,1 с –1 (± SE, n = 19) и 1,4 ± 0,1 с –1 (± SE, n = 18) и k медленный составляли 10,4 ± 0,3 с –1 (± SE, n = 19) и 9.9 ± 0,5 с –1 (± SE, n = 18) соответственно. Следовательно, не удалось обнаружить никакой разницы в скорости циклического электронного транспорта.
Пигментный состав
Состав неоксантина, лютеина, β-каротина, виолаксантина и антераксантина в растениях дикого типа и трансгенных растениях, лишенных PSI-H и PSI-L, показан в таблице 2. Отношение Chl a / b явно выше в двух трансгенных линий, чем у дикого типа (таблица 2). Эти значения хорошо согласуются со значениями, полученными для PSI-H спектрофотометрическими методами, и отражают тот факт, что растения, лишенные PSI-H или PSI-L, компенсируют это увеличением отношения PSI / PSII (Naver et al.1999). Содержание β-каротина не изменилось, но количество лютеина было значительно снижено (13–14%) у растений без PSI-H и PSI-L. Количество неоксантина также оказалось немного ниже (3%) у растений, лишенных PSI-H или PSI-L. Также была определена скорость образования антераксантина и зеаксантина во время обработки сильным светом. После 20 мин обработки высоким светом (500 мкмоль м –2 с –1 ) было замечено четкое изменение содержания виолаксантина, антераксантина и зеаксантина как у дикого типа, так и у растений без PSI-H и PSI-L. , что привело к ожидаемому изменению состояния эпоксидирования (EPS) (таблица 3).Не наблюдалось значительной разницы в количестве тушащих ксантофиллов или кинетике деэпоксидирования между растениями с дефицитом перехода состояний и диким типом.
Ультраструктура хлоропластов
Электронная микроскопия была проведена, чтобы выяснить, изменилась ли структура тилакоида в отсутствие PSI-H или PSI-L. Дикий тип образовывал хорошо выраженные граны, содержащие примерно 7,8 ± 1,2 стопки. Напротив, хлоропласты из растений без PSI-H и PSI-L образовывали более протяженные граны только с 4.4 ± 0,4 и 5,1 ± 0,6 стопки соответственно (± SE, n = 5, рис. 5). Количество стопок гран существенно различается ( P <10 –12 ).
Обсуждение
Неспособность выполнять переходы между состояниями не проявляется как визуальный фенотип
Чтобы определить, как неспособность выполнять переходы между состояниями влияет на высшее растение, растения Arabidopsis , лишенные PSI-H и PSI-L, были проанализированы в отношении роста и фотосинтетических характеристик в различных условиях освещения.Никаких различий в росте или морфологических характеристиках не было обнаружено ни в одном из тестируемых условий роста. Следовательно, растения могут компенсировать свой дефект как при слабом (30 мкмоль м –2 с –1 ), так и при высоком (400 мкмоль м –2 с –1 ) и очень сильном (1000 мкмоль м –2 с –1 ), оптимизируя свой фотосинтетический аппарат. У растений без PSI-H содержание PSI на 15% выше (Naver et al. 1999), и отсутствие переходов между состояниями, по-видимому, в значительной степени компенсируется более высоким содержанием PSI.Однако при стандартных условиях освещения в наших камерах для выращивания растения без PSI-H снизили фотохимическое тушение и эффективность PSII (Таблица 1). Таким образом, даже в условиях освещения, к которым растения адаптированы, отсутствие перехода между состояниями приводит к тому, что растения находятся в более неуравновешенном состоянии. Поскольку растения без PSI-H также имеют пониженную активность PSI, необходимо учитывать, влияет ли это на 1 – q P и ϕ PSII . У растений без PSI-N наблюдается аналогичное снижение активности PSI, однако у растений без PSI-N 1 – q P и ϕ PSII идентичны дикому типу в условиях освещения для роста (Haldrup et al.1999 г., данные не показаны). Чтобы предотвратить адаптацию к режиму постоянного освещения, также были проведены эксперименты по выращиванию при чередовании PSII / PSI или при слабом / высоком освещении. Тем не менее, даже в этих условиях не обнаруживались фенотипические различия между растениями дикого типа и растениями без PSI-H или PSI-L. Параметры флуоресценции, определенные для листьев, подвергшихся воздействию короткой серии чередующихся световых лучей PSI / PSII, показали, что растения дикого типа сохраняли почти постоянный уровень восстановления пула пластохинонов и эффективность фотохимии PSII при переходе из состояния 1 в состояние 2 (Таблица 1) .Напротив, растения с дефицитом PSI-H и PSI-L имели более низкое фотохимическое тушение под светом PSII, что отражало более уменьшенный пул пластохинона и более низкую эффективность PSII (Таблица 1). Параметры флуоресценции демонстрируют значительный дисбаланс в потоке электронов между двумя фотосистемами при отсутствии переходов между состояниями. Почему дисбаланс не приводит к визуальному фенотипу? Мутант ксантофилла ( npq1 ), лишенный виолаксантиндеэпоксидазы и, следовательно, неспособный превращать виолаксантин в антераксантин и зеаксантин, имеет резкое снижение как нефотохимического, так и фотохимического тушения, но также в этом случае нет очевидного фенотипа при различных условиях роста. (Niyogi et al.1998, Havaux and Niyogi 1999). У растений есть несколько механизмов защиты от стресса, и они, по-видимому, могут справиться с дефектами одной из систем светозащиты без серьезных последствий. Недавние результаты показали, что выращенные в полевых условиях мутанты npq4 , которые лишены энергозависимого тушения, дают на 40% меньше семян, чем дикий тип, хотя визуальный фенотип не наблюдался в камерах для роста при постоянных условиях (Külheim et al. 2002). Таким образом, реальное значение белка может стать очевидным только в изменяющихся условиях.При чередовании света PSI / PSII не было различий между растениями дикого типа и растениями без PSI-H в производстве семян, однако при чередовании высокого / низкого освещения растения без PSI-H показали 10% снижение количества растений. набор семян на растение. Как обсуждается ниже, общая фотосинтетическая эффективность снижается на 8–14%, поэтому можно ожидать аналогичного снижения приспособленности. Хотя такое незначительное изменение приспособленности имело бы сильное влияние на выживание видов растений в природе, его трудно определить с уверенностью в экспериментах по контролируемому росту.Из-за естественного большого разброса количества семян, заложенных на одно растение, необходимо проанализировать приблизительно 800 растений дикого типа и растений без PSI-H, чтобы определить, является ли наблюдаемое снижение статистически значимым.
Снижение фотосинтетической способности
Растения без PSI-H, которые были акклиматизированы к PFD 100 мкмоль / м –2 с –1 показали как снижение квантового выхода на 8%, так и снижение максимальной фотосинтетической способности на 14% при ярком освещении (рис.3). Однако при интенсивности света для роста растения, лишенные PSI-H, работали так же хорошо, как и растения дикого типа. Для растений без PSI-L наблюдалось аналогичное, но менее серьезное снижение квантового выхода и эффект максимальной фотосинтетической способности. Таким образом, растения с недостаточным переходом между состояниями способны почти полностью компенсировать рост на свету, но они демонстрируют ограниченный динамический диапазон по сравнению с диким типом. Более низкая максимальная фотосинтетическая способность и более низкая точка компенсации света являются особенностями, напоминающими теневые листья, и было возможно, что растения без PSI-H имели другие характеристики теневых листьев e.грамм. более низкое содержание ферментов цикла Кальвина. Корреляция между максимальной скоростью фотосинтеза и концентрацией Rubisco ранее была показана на растениях Arabidopsis , акклиматизированных к постоянному высокому или низкому освещению (Walters and Horton 1994, Stitt and Schulze 1994). Однако не было обнаружено значительного изменения количества Rubisco в растениях без PSI-H (данные не показаны).
Без компенсации с помощью альтернативных механизмов закалки
Ксантофилловый цикл является основным способом избавления от избыточной энергии возбуждения во время светового стресса, но также и от других пигментов e.грамм. лютеин, по-видимому, участвует (обзор в Niyogi 1999, Niyogi et al. 2001). Поэтому мы рассмотрели, будут ли растения без PSI-H и PSI-L реагировать на увеличение (1 – q P ) через этот альтернативный механизм тушения. Однако содержание виолаксантина или лютеина не увеличивалось в адаптированных к темноте листьях (таблица 2). Фактически, было небольшое уменьшение этих пигментов по сравнению с хлорофиллом из-за увеличения содержания PSI. Аналогичное снижение наблюдалось и в неоксантине, который обнаружен только в связке с Lhcb1-5 (Bassi and Caffarri 2000).Мы также считали, что активность виолаксантин деэпоксидазы, фермента, который превращает виолаксантин в антераксантин и зеаксантин при ярком освещении, была повышена, но не было обнаружено никакой разницы в скорости деэпоксидирования (таблица 3). В заключение, отсутствие переходов между состояниями не вызывает заметных изменений в цикле ксантофилла.
Малатный клапан, закрывающий преобразование оксалоацетата в малат, действует как акцептор электронов от PSI, когда фотосистемы находятся в опасности чрезмерного восстановления (Backhausen et al.2000). Линейный перенос электронов к оксалоацетату катализируется NADP + -малатдегидрогеназой, и, следовательно, емкость малатного клапана зависит от активности этого фермента. Активность фермента была идентична у растений с дефицитом перехода между состояниями и у растений дикого типа, поэтому маловероятно, что большее количество электронов ускользает по этому пути (рис. 4). Альтернативными путями фотозащиты может быть прямое восстановление O 2 с помощью PSI (Asada 1999) или циклический перенос электронов (Niyogi 1999).Однако не было обнаружено никаких указаний на то, что растения, лишенные PSI-H, отличались от дикого типа по этим путям.
Изменения ультраструктуры тилакоидов
У тилакоидов растений, лишенных PSI-H или PSI-L, наблюдается значительно измененная ультраструктура. Грана выглядела более протяженной и содержала меньше дисков на грану (рис. 5). Аналогичное изменение ультраструктуры наблюдалось у нескольких видов растений, выращенных под светом ФС II (Deng et al. 1989, Lui et al.1993). Эти растения показали, как и растения без PSI-H и PSI-L, более высокое соотношение Chl a / b и повышенное соотношение PSI / PSII (Naver et al. 1999). Таким образом, возможным объяснением меньшего количества укрупненных граней может быть то, что повышенная концентрация PSI препятствует образованию гран. Однако растения без PSI-N также содержат на 20% больше PSI, чем растения дикого типа, но не обнаруживают соответствующих изменений в ультраструктуре хлоропластов (Haldrup et al. 1999, неопубликованные результаты A. Haldrup).Могут ли изменения в организации LHCII быть ответственными за измененную структуру тилакоидов? Участие LHCII в накоплении граны до сих пор остается спорным. Включение LHCII в липидные везикулы в присутствии Mg + приводит к образованию мембранных аппрессий, а мягкая обработка трипсином для удаления открытых частей белка полностью устраняет образование стопок (Mullet and Arntzen 1980). Это указывает на то, что трехмерные взаимодействия LHCII важны для стэкинга.Однако мутантные растения ячменя, лишенные LHCII, все еще способны образовывать стопки гран (Simpson 1979). Было показано, что растения без PSI-H имеют сильно фосфорилированный LHCII по сравнению с диким типом (Lunde et al. 2001, Haldrup et al. 2001). Более высокий уровень фосфорилирования обнаруживается во всех тестируемых условиях (высокий, низкий, рост, PSI- или PSII-свет) и даже после длительной адаптации к темноте. Было высказано предположение, что фосфорилирование антенных белков PSII мешает упаковке белка и тем самым препятствует образованию четко выраженных гран (Staehelin and Arntzen 1983, Georgakopoulos and Argyroudi-Akoyunoglou 1994).В подтверждение этой модели разупаковка наблюдалась при переходе из состояния 1 в состояние 2 у водорослей и видов растений, включая Arabidopsis (Barber 1982, Rozak et al. 2002, неопубликованные результаты A. Haldrup). Сходный эффект на структуру тилакоидов наблюдался у растений, лишенных PSI-F или PSI-E, где уровень фосфорилирования LHCII также значительно повышен (Haldrup et al. 2000, Pesaresi et al. 2002). Характеристика белковых взаимодействий между слоями в гранах показала, что ФСII во многих случаях сталкивается со слоем, состоящим только из LHCII (Boekema et al.2000). Таким образом, возможно, что фосфорилирование LHCII напрямую нарушает 3-мерное взаимодействие между PSII и LHCII и, таким образом, плотную упаковку стопок гран (Bennett 1983, Allen 1992).
Если уменьшение укладки гранов отражает измененную молекулярную организацию LHCII, может ли это объяснить отсутствие визуального фенотипа в условиях высокой освещенности? Судьба поглощенного света зависит от молекулярной организации LHCII, формируя либо светособирающее, либо рассеивающее энергию взаимодействие (Horton 1999, Garab and Mustárdy 1999, Allen and Forsberg 2001).Когда LHCII упорядочен в грана, светособирающая форма преобладает, позволяя переносить энергию от монослоя LHCII к комплексам PSII в противоположном слое (Boekema et al. 2000). Однако после разложения LHCII становится дезорганизованным и вместо этого действует как тушитель. Кривые индукции флуоресценции показали, что в присутствии АТФ требуется больше Mg 2+ для ослабления тушения (Horton and Black 1983). Это указывает на то, что фосфорилирование способствует диссипативной форме LHCII.Следовательно, возможно, что часть LHCII в растениях с дефицитом перехода между состояниями существует в форме, благоприятствующей тушению, и что это в некоторой степени защищает растения в других условиях фотоингибирования. Ожидаемая стоимость наличия таких гасящих агрегатов LHCII будет заключаться в более низкой фотосинтетической эффективности, как это наблюдается у растений без PSI-H и PSI-L.
Заключение
В нормальных условиях роста растения, у которых отсутствует переход между состояниями, имеют более уменьшенный пул пластохинона и более низкую активность ФСII.В условиях состояния 2 они не могут активировать нефотохимическое тушение. Напротив, дикий тип способен оптимизировать свои фотосинтетические характеристики при всех протестированных световых режимах и поддерживать постоянными квантовый выход и (1 – q P ). В отсутствие переходов между состояниями эффективность использования света снижается примерно на 10%, но эта уменьшенная фотосинтетическая способность не приводит к визуальному фенотипу ни в одном из тестируемых условий роста. Мы предполагаем, что отсутствие переходов между состояниями в значительной степени может быть нейтрализовано увеличением отношения PSI / PSII и уменьшением организации LHCII в стеках гран.Данные показывают, что переходы между состояниями не нужны ни для оптимизации фотосинтеза при слабом освещении, ни для фотозащиты при ярком освещении. Вместо этого переходы между состояниями, по-видимому, являются механизмом тонкой настройки для увеличения динамического диапазона света, в котором фотосинтез является оптимальным, что играет важную роль в среде с изменяющимися условиями освещения.
Материалы и методы
Растительный материал, иммуноблот-анализ и выделение тилакоидных мембран
Arabidopsis thaliana Heynh.экотип Columbia и его трансформанты выращивали в камерах Arabidopsis (Percival AR-60 I, Бун, Айова, США). Стандартными условиями были, если не указано иное, 20 ° C, влажность 70%, 100–120 мкмоль м –2 с –1 и 8-часовой фотопериод. Эксперимент с различными условиями освещения проводился в камере, оснащенной ртутными лампами NQI. Температура, влажность и фотопериод составляли 12 или 8 часов. Свет PSII был получен путем фильтрации света с помощью синего фильтра (183 Moonlight Blue, Lee Filters, Andover, Hants, U.K.) и свет PSI путем фильтрации с красным фильтром (026 Medium Red, Lee Filters). Аналогичное фотосинтетически активное излучение (40 мкмоль м –2 с –1 ) при ФСII и ФСI было получено с использованием серых фильтров (209 Neutral Gray, Lee Filters). Тот же нейтральный фильтр использовали для выращивания при переменной интенсивности света (40 и 600 мкмоль м –2 с –1 ). Растения, лишенные PSI-H, были получены, как описано ранее (Naver et al. 1999). Растения, лишенные PSI-L, были получены путем клонирования частичного клона кДНК [инвентарный номер AT50519 (23002T7)] в антисмысловой ориентации между усиленным промотором 35S CaMV и терминатором 35S.Векторы и процедуры трансформации были такими же, как у Jensen et al. (2002). Растения, лишенные PSI-H и -L, идентифицировали путем приготовления экстрактов цельного листа, разделения белков с помощью SDS-PAGE и иммуноблоттинга с использованием специфических антител к PSI-H и -L.
Тилакоидные мембраны очищали, как описано ранее (Haldrup et al. 1999). Иммуноблоты выполняли по стандартным методикам. Все значения P для статистической значимости были определены с использованием двустороннего критерия Стьюдента t .
Измерения флуоресценции и кислорода
Параметры флуоресценции (NPQ, q P и ϕ PSII ) при различной интенсивности света получали, как описано ранее (Haldrup et al. 1999). Максимальный квантовый выход ФСII (ϕ PSII ), фотохимическое (q P ) и нефотохимическое тушение (NPQ) были рассчитаны как ϕ PSII = (F m ′ –F S ) / F м ′, q P = (F m ′ –F s ) / (F m ′ –F 0 ′), и NPQ = (F m –F m ′) / F m ′, соответственно, где F S — стационарная флуоресценция, F m и F m ′ — максимальная флуоресценция, индуцированная насыщающей вспышкой длительностью 800 мс (6000 мкмоль m -2 s –1 ) адаптированных к темноте или предварительно освещенных листьев, а F 0 и F 0 ′ — флуоресценция без освещения с актиничным светом адаптированных к темноте и предварительно освещенных листьев (Anderson et al.1997). Параметры флуоресценции при переменном освещении PSI / PSII определяли на отдельных листьях с помощью флуориметра с амплитудно-импульсной модуляцией (PAM) 101–102–103 (Walz, Effeltrich, Германия). Лист подвергался воздействию фотонов 100 мкмоль –2 с –1 синего света от лампы Schott KL-1500, оснащенной фильтром Corning 4–96, и с интервалом в 15 минут дальним красным светом (Waltz 102-FR ) был включен или выключен. Максимальный выход флуоресценции в состояниях 1 и 2 определяли по насыщающей вспышке белого света длительностью 800 мс (6000 мкмоль м –2 с –1 ).
Скорость выделения кислорода определялась, как описано ранее (Haldrup et al. 1999). Площадь листа определяли с использованием BioRad Gel Doc 2000, BioRad Quantity One 4.0.2. программное обеспечение. Для статистического анализа определяли квантовый выход, точку компенсации и максимальное выделение кислорода для каждого из отдельных растений, и сравнения проводили с использованием теста Стьюдента t . Отражение света определяли сканированием (Epson 1600 Pro) листьев и количественной оценки отраженного света путем сравнения с эталоном шкалы серого Kodak (Kodak, Торонто, Канада).Прохождение света через лист определяли путем измерения интенсивности света (Schott KL-1500) от световода до и после того, как лист был помещен между волокном и квантовым датчиком (Li-Cor, Quantum, США).
Циклический перенос электронов
Циклический перенос электронов был определен путем измерения индуцированных светом изменений поглощения P700 на длине волны 810–860 нм с использованием двухволнового эмиттерного детектора Walz ED-P700DW-E, подключенного через флуорометр PAM к осциллографу Tektronix TDS420.Использовали листья адаптированных к темноте растений. P700 был возбужден путем освещения дальним красным светом 735 ± 40 нм от Walz 102-FR. За индуцированными изменениями поглощения P700 наблюдали в течение 20 с, актиничный свет отключили и повторное восстановление P700 + регистрировали еще в течение 25 с. Для каждого листа усредняли четыре следа. Циклический перенос электронов оценивался по повторному восстановлению P700 + , то есть уменьшению поглощения P700 после выключения актиничного света.Постиллюминационное изменение оптической плотности было преобразовано в экспоненциальное затухание с помощью нелинейной регрессии Левенберга-Марквардта.
Пигментный анализ
Для анализа пигмента полностью сформировавшиеся листья собирали после 16 часов адаптации к темноте. Затем растения освещали фотонами 500 мкмоль –2 с –1 , и листья собирали с каждого растения через 15, 30, 60 и 120 минут, немедленно замораживали в жидком азоте и хранили при –80 ° C до анализ.Все экстракции производились при тусклом зеленом свете. Пигменты экстрагировали, как описано в García-Plazaola and Becerril (1999). Состав пигмента определяли, как в Jensen et al. (2002).
НАДФ
+ -фотовосстановление и анализ НАДФ-малатдегидрогеназыНАДФ + -фотовосстановление in vitro определяли по изменениям оптической плотности при 340 нм, по существу, как описано ранее (Kj¾r and Scheller 1996, Naver et al. 1996).В анаэробных условиях образцы промывали аргоном и реакцию проводили в присутствии 4 мкг каталазы, 6 мМ глюкозы и 2 единиц глюкозооксидазы (Kj¾r and Scheller 1996). Экстракцию листьев и измерения активности НАДФ-малатдегидрогеназы проводили, в основном, как у Scheibe и Stitt (1988).
Электронная микроскопия
Электронную микроскопию выполняли, как описано в Haldrup et al. (2000).
Благодарность
Д-р Дэвид Дж.Благодарим Симпсона за его помощь с электронной микроскопией, доктора Стефана Янссона за предоставление антител против белков Lhca и профессора Биргера Линдберга Мёллера за ценные обсуждения. Работа поддержана Датским национальным исследовательским фондом.
Рис. 1 Иммуноблот-анализ накопления субъединиц PSI и светособирающих белков в антисмысловых PsaL и растениях дикого типа. (A) Белки, эквивалентные 1 мкг хлорофилла, были загружены для обнаружения субъединиц PSI и 0.0625–0,125 мкг для обнаружения светособирающих белков. Моноклональное антитело против Lhca2 также распознает Lhcb4. (B) Для количественного определения остаточных количеств PSI-L и PSI-H в растениях без PSI-L, 3 мкг хлорофилла были загружены в WT и -L дорожку, и серию двукратных разведений WT на дорожке От 0,5 × WT до 0,015 × WT.
Рис. 1 Иммуноблот-анализ накопления субъединиц PSI и светособирающих белков в антисмысловых PsaL и растениях дикого типа.(A) Белки, эквивалентные 1 мкг хлорофилла, загружали для обнаружения субъединиц PSI и 0,0625–0,125 мкг для обнаружения светособирающих белков. Моноклональное антитело против Lhca2 также распознает Lhcb4. (B) Для количественного определения остаточных количеств PSI-L и PSI-H в растениях без PSI-L, 3 мкг хлорофилла были загружены в WT и -L дорожку, и серию двукратных разведений WT на дорожке От 0,5 × WT до 0,015 × WT.
Рис. 2 Зависимость от света параметров флуоресценции хлорофилла у дикого типа и листьев без PSI-H.(A) Нефотохимическое тушение, NPQ. (B) Квантовый выход фотохимии ФСII, ϕ ФСII . (C) Фотохимическое тушение 1-q P , представляющее оценку состояния восстановления Q A . Каждый график представляет собой среднее значение шести растений, измеренных на трех разных листьях. Полоски указывают стандартные ошибки.
Рис. 2 Зависимость от света параметров флуоресценции хлорофилла у дикого типа и листьев без PSI-H. (A) Нефотохимическое тушение, NPQ.(B) Квантовый выход фотохимии ФСII, ϕ ФСII . (C) Фотохимическое тушение 1-q P , представляющее оценку состояния восстановления Q A . Каждый график представляет собой среднее значение шести растений, измеренных на трех разных листьях. Полоски указывают стандартные ошибки.
Рис. 3 Кривая светового отклика для выделения кислорода у дикого типа и листьев, лишенных PSI-H (A). Квантовый выход для дикого типа и растений без PSI-H определяли по наклону линейной части кривой при слабом актиничном свете (B).Кривые представляют собой среднее значение для шести растений, каждое из которых измерено трижды. Полоски указывают стандартные ошибки. Было определено, что поглощение света листьями дикого типа и без PSI-H составляет 0,80 и 0,79 соответственно (SE ± 0,02).
Рис. 3 Кривая светового отклика для выделения кислорода у дикого типа и листьев, лишенных PSI-H (A). Квантовый выход для дикого типа и растений без PSI-H определяли по наклону линейной части кривой при слабом актиничном свете (B). Кривые представляют собой среднее значение для шести растений, каждое из которых измерено трижды.Полоски указывают стандартные ошибки. Было определено, что поглощение света листьями дикого типа и без PSI-H составляет 0,80 и 0,79 соответственно (SE ± 0,02).
Рис. 4 Уровень активации НАДФ + -малатдегидрогеназы. Листья собирали через 15 мин, 2 ч и 4 ч после появления света в камере для выращивания. Общую активность получали путем воздействия на белковый экстракт в течение 40 мин 0,25 M DTT при pH 9 перед измерением.
Фиг.4 Уровень активации НАДФ + -малатдегидрогеназы. Листья собирали через 15 мин, 2 ч и 4 ч после появления света в камере для выращивания. Общую активность получали путем воздействия на белковый экстракт в течение 40 мин 0,25 M DTT при pH 9 перед измерением.
Рис. 5 Ультраструктура хлоропластов (A) дикого типа, (B) PSI-H и (C) без PSI-L, суспендированных в 10 мМ MgCl 2 , 0,06 мМ натрий-фосфатном буфере (pH 7,4) и 2% глутарового альдегида.
Рис. 5 Ультраструктура (A) хлоропластов дикого типа (B) PSI-H и (C) PSI-L-без хлоропластов, суспендированных в 10 мМ MgCl 2 , 0,06 мМ натрий-фосфатном буфере (pH 7,4) и 2% глутарового альдегида.
Таблица 1 Давление возбужденияPSII (1-q P ), квантовый выход фотохимии PSII (ϕ PSII ) и нефотохимическое тушение (NPQ), измеренные в процессе роста, PSI и свет PSI
| Свет | Завод | (1 – q P ) | ϕ PSII | NPQ | |||
| Лампа для выращивания | Дикий тип | 0.008 ± 0,002 | 0,75 ± 0,01 | nd a | |||
| Нет PSI-L | nd | nd | nd | ||||
| нет | |||||||
| 0,0 | *0,70 ± 0,01 * | nd | |||||
| PSI (состояние 1) | Дикий тип | 0,11 ± 0,02 | 0,69 ± 0,02 | 0,21 ± 0,02 | |||
| L | — 0.15 ± 0,010,67 ± 0,01 | 0,19 ± 0,01 | |||||
| Нет PSI-H | 0,15 ± 0,01 | 0,66 ± 0,01 | 0,15 ± 0,02 | ||||
| PS тип | 0,13 ± 0,01 | 0,66 ± 0,01 | 0,29 ± 0,02 | ||||
| Нет PSI-L | 0,22 ± 0,01 * | 0,57 ± 0,01 * | 0,19 ± 0,0302 | Нет PSI-H0.26 ± 0,01 * | 0,55 ± 0,01 * | 0,16 ± 0,02 * |
| Легкий | Завод | (1 – кв. | Лампа для выращивания | Дикий тип | 0,008 ± 0,002 | 0,75 ± 0,01 | nd a | ||
| Нет PSI-L | nd | nd6 | nd | Нет PSI-H | 0.082 ± 0,004 * | 0,70 ± 0,01 * | nd | ||
| PSI (состояние 1) | Дикий тип | 0,11 ± 0,02 | 0,69 ± 0,02 | 0,21 ± 0,02 | |||||
| L | 0,15 ± 0,01 | 0,67 ± 0,01 | 0,19 ± 0,01 | ||||||
| Нет PSI-H | 0,15 ± 0,01 | 0,66 ± 0,01 | 0,15 ± 0,02 | ||||||
| (состояние | ) | Дикий тип | 0.13 ± 0,01 | 0,66 ± 0,01 | 0,29 ± 0,02 | ||||
| Нет PSI-L | 0,22 ± 0,01 * | 0,57 ± 0,01 * | 0,19 ± 0,01 * | ||||||
| H | 0,26 ± 0,01 * | 0,55 ± 0,01 * | 0,16 ± 0,02 * |
Давление возбуждения ФСII (1-q P ), квантовый выход фотохимии ФСII (ϕ13412 ФСIIII ) и нефотохимическое тушение (NPQ), измеренное в условиях роста, PSI и PSII light
| Light | Plant | (1 – q P ) | ϕ PSII | NPQ | Growth светлый | Дикий тип | 0.008 ± 0,002 | 0,75 ± 0,01 | nd a | |
| Нет PSI-L | nd | nd | nd | |||||||
| нет | ||||||||||
| 0,0 | *0,70 ± 0,01 * | nd | ||||||||
| PSI (состояние 1) | Дикий тип | 0,11 ± 0,02 | 0,69 ± 0,02 | 0,21 ± 0,02 | ||||||
| L | — 0.15 ± 0,010,67 ± 0,01 | 0,19 ± 0,01 | ||||||||
| Нет PSI-H | 0,15 ± 0,01 | 0,66 ± 0,01 | 0,15 ± 0,02 | |||||||
| PS тип | 0,13 ± 0,01 | 0,66 ± 0,01 | 0,29 ± 0,02 | |||||||
| Нет PSI-L | 0,22 ± 0,01 * | 0,57 ± 0,01 * | 0,19 ± 0,0302 | Нет PSI-H0.26 ± 0,01 * | 0,55 ± 0,01 * | 0,16 ± 0,02 * |
| Легкий | Завод | (1 – кв. | Лампа для выращивания | Дикий тип | 0,008 ± 0,002 | 0,75 ± 0,01 | nd a | ||
| Нет PSI-L | nd | nd6 | nd | Нет PSI-H | 0.082 ± 0,004 * | 0,70 ± 0,01 * | nd | ||
| PSI (состояние 1) | Дикий тип | 0,11 ± 0,02 | 0,69 ± 0,02 | 0,21 ± 0,02 | |||||
| L | 0,15 ± 0,01 | 0,67 ± 0,01 | 0,19 ± 0,01 | ||||||
| Нет PSI-H | 0,15 ± 0,01 | 0,66 ± 0,01 | 0,15 ± 0,02 | ||||||
| (состояние | ) | Дикий тип | 0.13 ± 0,01 | 0,66 ± 0,01 | 0,29 ± 0,02 | ||||
| Нет PSI-L | 0,22 ± 0,01 * | 0,57 ± 0,01 * | 0,19 ± 0,01 * | ||||||
| H | 0,26 ± 0,01 * | 0,55 ± 0,01 * | 0,16 ± 0,02 * |
Состав пигментов у растений дикого типа и растений без PSI-H или PSI-L
| Пигмент | Дикий тип | PSI-L меньше | PSI-H меньше |
| Chl a / b | 2.48 ± 0,03 | 2,78 ± 0,08 * | 2,60 ± 0,03 * |
| Неоксантин | 6,1 ± 0,5 | 5,9 ± 0,5 | 5,9 ± 0,3 |
| Лютеин | 2,117302 17 * | 15,3 ± 1,3 * | |
| β-каротин | 15,6 ± 1 | 16,0 ± 1 | 15,3 ± 0,5 |
| Виолаксантин (D) | 5,4 ± 1 | 5,1 ± 15,1 ± 0,5 | |
| Антераксантин (D) | 0.4 ± 0,2 | 0,4 ± 0,2 | 0,5 ± 0,1 |
| Пигмент | Дикий тип | PSI-L меньше | PSI-H меньше |
| Chl b a | 2,48 ± 0,03 | 2,78 ± 0,08 * | 2,60 ± 0,03 * |
| Неоксантин | 6,1 ± 0,5 | 5,9 ± 0,5 | 5,9 ± 0,3 |
| 15302 | |||
| .4 ± 1,6 * | 15,3 ± 1,3 * | ||
| β-каротин | 15,6 ± 1 | 16,0 ± 1 | 15,3 ± 0,5 |
| Виолаксантин (D) | 5,4 ± 1 | 5,2 ± 0,5 | |
| Антераксантин (D) | 0,4 ± 0,2 | 0,4 ± 0,2 | 0,5 ± 0,1 |
Состав пигментов у растений дикого типа и растений, лишенных PSI -H или PSI-L
| Пигмент | Дикий тип | PSI-L меньше | PSI-H меньше |
| Chl a / b | 2.48 ± 0,03 | 2,78 ± 0,08 * | 2,60 ± 0,03 * |
| Неоксантин | 6,1 ± 0,5 | 5,9 ± 0,5 | 5,9 ± 0,3 |
| Лютеин | 2,117302 17 * | 15,3 ± 1,3 * | |
| β-каротин | 15,6 ± 1 | 16,0 ± 1 | 15,3 ± 0,5 |
| Виолаксантин (D) | 5,4 ± 1 | 5,1 ± 15,1 ± 0,5 | |
| Антераксантин (D) | 0.4 ± 0,2 | 0,4 ± 0,2 | 0,5 ± 0,1 |
| Пигмент | Дикий тип | PSI-L меньше | PSI-H меньше |
| Chl b a | 2,48 ± 0,03 | 2,78 ± 0,08 * | 2,60 ± 0,03 * |
| Неоксантин | 6,1 ± 0,5 | 5,9 ± 0,5 | 5,9 ± 0,3 |
| 15302 | |||
| .4 ± 1,6 * | 15,3 ± 1,3 * | ||
| β-каротин | 15,6 ± 1 | 16,0 ± 1 | 15,3 ± 0,5 |
| Виолаксантин (D) | 5,4 ± 1 | 5,2 ± 0,5 | |
| Антераксантин (D) | 0,4 ± 0,2 | 0,4 ± 0,2 | 0,5 ± 0,1 |
Состояние эпоксидирования ксантофиллов во время обработки высоким светом у растений дикого типа и растений, лишенных PSI-H или PSI-L